Original Article
Anatomical Particularities of the Mammary Glands in the Adult Female Grasscutter (Thryonomys swinderianus, Temminck, 1827)
|
Gualbert
Simon Nteme Ella 1* 1 Department of Anatomy-Histology-Embryology, Inter-State School of
Veterinary Sciences and Medicine (EISMV) of Dakar, Senegal 2 Institute
for Public Law Studies (IEDP), Jean Monnet Faculty of the University of
Paris-Saclay, France 3 Department of Biology and Animal Production-Nangui University Abrogoua. 02
BP: 801 Abidjan 02, Côte D’Ivoire 4 Institut National Des Recherches Agricoles Du Benin –
CRA-Agonkanmey 01 884 Recette
Principale, Cotonou 01, Benin |
|
|
|
ABSTRACT |
||
|
This study aimed to characterize the mammary apparatus of the adult female grasscutter, or aulacode (Thryonomys swinderianus Temminck, 1827), by providing a detailed topographical description of the mammary anatomy in five adult, farm-raised females with an average body weight of 3.5 kg. Examination of the mammary gland revealed anatomical features that are broadly comparable to those observed in other rodent species and even in lagomorphs; however, several specific features were observed in this species. Three pairs of mammary papillae, which are relatively short but noticeably broader than those of commonly described domestic rodents, were observed in the pectoral, thoracic, and abdominal regions, respectively. Subcutaneously, the mammary glands on each flank are fused into a continuous mass of glandular tissue extending from the pectoral region to the inguinal region. Longitudinally—that is, from the pectoral to the inguinal region—the length of each mass averages 21.5 ± 3.5 cm. As for the lateromedial extent, that is, the width of this glandular mass, it averages 6.5 ± 1.5 cm. This structural configuration represents a distinctive characteristic of the species when compared with domestic rodents and lagomorphs that exhibit similar litter sizes. The biomorphometric features described here are likely to influence maternal behavior in captive female grasscutters and should support improved anatomophysiological management of reproduction in this characteristically African rodent. This is particularly relevant for the implementation of cross-fostering strategies in grasscutter farming, aimed at managing litter sizes and adjusting the weight of young, especially in cases of lactation difficulties or hyperprolific litters. Keywords: Grasscuter (Thryonomys Swinderianus),
Mammary Gland, Topography, Conformation, Reproductive Characteristics. |
||
INTRODUCTION
The grasscutter,
or aulacode (Thryonomys swinderianus
Temminck, 1827), is a wild hystricomorph rodent whose intensive captive
breeding is increasingly expanding in sub-Saharan Africa for food production
and wildlife management. In parts of Central and West Africa, where grasses
form their natural habitat and primary food source, these rodents are commonly
referred to as “grasscutters.” In South Africa,
however, they are associated with cane plantations, where they are known as
cane rats and regarded as significant agricultural pests. Accordingly, the
distribution of the greater cane rat is largely determined by the availability
of, or preference for, particular grass species as a food source (Baptist and
Mensah, 1986; Byanet et al., 2009).
The meat of this
rodent, which is not subject to any cultural taboos, is highly valued by
African consumers of bushmeat. Numerous studies have been conducted on the aulacode to improve breeding techniques by enhancing
understanding of its reproduction, growth, and behavior
Ewer (1969), Asibey (1974), Adjanohoun
(1992), Tondji and
Agbessi (1992), Yewadan (1992), Houben
(1999), Fantodji and
Soro (2004), Abe (2010), Broalet et al.
(2012). However, very few investigations to date have focused specifically on
the reproductive characteristics of this typically African rodent during the
process of domestication Adjanohoun
(1992), Addo et al. (2007), Soro (2007).
To address the
scarcity of anatomical data on aulacode reproduction,
we conducted a descriptive study of the mammary glands in adult females. In
pubertal females, the glands are functional, producing colostrum and milk after
parturition. Colostrum is a nutritive fluid for the neonate, rich in maternal
immunoglobulins that confer passive humoral immunity. Milk is a complete
nutritional source capable of fully meeting the offspring’s needs and remains
the sole source of nourishment until weaning. Among all females, the mammary
glands are not identical; they may differ in their position along the ventral
surface of the trunk and in their anatomical characteristics Raynaud
(1961), Grassé (1971), Hoshino
(1979), Hovey
and Trott (2004), Vallantin
(2023). This is essential, as anatomy underpins medical and surgical
practice. Compared with domestic species, the grasscutter is relatively
disease-resistant due to its natural hardiness; however, captive individuals
are more susceptible to dystocia or other birthing difficulties Jori et al. (2001); Fantodji and
Soro (2004), Houben
et al. (2004), Yapi et al. (2013), Vétérinaires Sans Frontières. (n.d.). Consequently, improved management of this
species depends on a deeper understanding of its morphology, physiology, and
behaviour. In particular, morphological studies require the integration of
multiple anatomical disciplines, including general, descriptive, topographical,
functional, developmental, and comparative anatomy.
This study
primarily aimed to advance knowledge of grasscutter anatomy, with particular
emphasis on providing a detailed topographical description of the adult female
mammary apparatus. It also provides baseline information to assist farmers and
researchers in promoting sustainable grasscutter farming and reducing reliance
on wild populations.
Materials and methods
Materials
The grasscutter
(Thryonomys swinderianus Temminck, 1827) is currently
listed as Least Concern (LC) on the International Union for Conservation of
Nature (IUCN) Red List Child
(2016).. In this context, the comparative anatomy
collection of the Department of Anatomy, Histology, and Embryology at the Ecole
Inter-Etats des Sciences et Médecine
Vétérinaires (EISMV) in Dakar is sourced from two
main origins. The primary source, representing approximately two-thirds of the
specimens, consists of tissues obtained from carcasses of animals slaughtered
in establishments specializing in the preparation and sale of grasscutter meat
for human consumption. The secondary, less frequent source comprises specimens
from animals that died suddenly in captivity or were euthanized for medical
(e.g., poor health, severe suffering) or sanitary reasons. It should be noted
that, in addition to Senegal, the animals included in this comparative anatomy
collection originate from farms in various countries of the sub-region, such as
Abidjan (Côte d’Ivoire) and Cotonou (Benin). When necessary, and to facilitate
their transport to Dakar, the carcasses are exsanguinated and either
refrigerated or frozen to minimize the degradation of anatomical structures.
In addition to
facilitating comparative studies of anatomical structures across different
vertebrate groups, this collection—established at the founding of EISMV—allows
the study of various animal species without the need for additional sampling
from the wild. In this context, the animals used in the present study were
adult females that had completed their reproductive cycle and were therefore
retired from breeding and sold as grasscutter meat for human consumption.
Specifically, five adult female grasscutters with an
average live weight of 3.5 kg, obtained from intensive captive breeding
facilities (grasscutter breeding), were used. The dissection of the entire
mammary apparatus was performed using small surgical instruments suitable for
laboratory animals.
Ethical Considerations and Experimental Procedures
The EISMV is
affiliated with Cheikh Anta Diop University (UCAD) in Dakar. Accordingly, all
experimental procedures were carried out in compliance with the recommendations
of the UCAD Research Ethics Committee UCAD (2024), the provisions of the European Convention
for the Protection of Vertebrate Animals used for Experimental and Other
Scientific Purposes (1986), the ARRIVE guidelines (Animal Research: Reporting
of In Vivo Experiments), and applicable Senegalese legislation on animal
welfare.
Where applicable,
the dissection and mammary gland visualization techniques followed methods
described by previous authors Popesko (1972), Barone
(1976), Bourdelle and Bressou,
1978, Dyce et al. (1996, which are standard for laboratory animals.
For each animal, five procedural steps were performed: immobilization and
restraint (manual for docile individuals or using a cage for non-docile
subjects); sedation and anesthesia with ketamine (Imalgène ND) at a dose of 0.7 mg/kg, administered via the
external jugular vein located in the jugular groove; exsanguination; cleaning;
and, finally, skinning. Prior to skinning, with the animal positioned in dorsal
recumbency, a topographical description of the mammary chain was performed over
the thoraco-abdomino-pelvic region. Thereafter, the
animal was skinned using a scalpel and mouse-tooth forceps to fully expose the
mammary apparatus. The mammary glands were subsequently examined and described
in accordance with models established for anatomical studies in other animal
species Barone
et al. (1973), Lebas
(2002). The anatomical characteristics recorded focused primarily on the
topography, conformation, and anatomical relationships of the mammary glands.
Results
Regarding the
topography of the mammary glands, in female grasscutters positioned in dorsal
recumbency, small pinkish nodules were observed on the ventral region of the
trunk beneath the white coat, which is sparsely covered with subspiny hair. These nodules correspond to the mammary
papillae or teats, arranged on either side of the midline in three rows,
corresponding to three pairs of mammary glands. Specifically, beneath the hair,
the mammary glands are located in the pectoral, thoracic, and abdominal regions
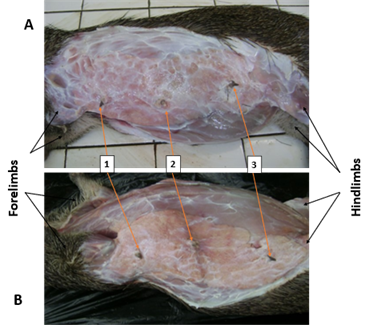
Figure 1.
|
Figure 1
|
|
Figure 1 Subcutaneous Topography of the
Mammary Gland of the Female Grasscutter: A- Right Lateral Recumbency; B- Left
lateral recumbency; 1- Pectoral teat; 2- Thoracic teat; 3- Abdominal teat |
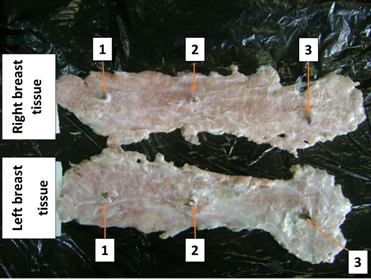
To examine the
morphology of the mammary glands Figure 1 and Figure 2, the ventral region of the trunk was skinned to expose the glands.
Each mammary gland was observed to consist of two elongated, thick layers of
glandular tissue arranged parallel to the linea alba.
Consequently, the mammary glands on each flank are fused into a continuous mass
of glandular tissue extending from the pectoral region to the inguinal region.
Longitudinally—that is, from the pectoral to the inguinal region—the length of
each mass averages 21.5 ± 3.5 cm. Regarding the lateromedial extent, that is,
the width of this glandular mass, it averages 6.5 ± 1.5 cm.
|
Figure 2
|
|
Figure 2 External Conformation – Continuous
Mass of Mammary Glandular Tissue in the Female Grasscutter: Ventral View of
Isolated Mammary Glands. 1- Pectoral teat; 2- Thoracic teat; 3- Abdominal
teat |
Discussion
The grasscutter
trade remains far less developed and structured than that of cattle, sheep, or
poultry Fantodji and
Soro (2004), Houben
et al. (2004), Yapi et al. (2013), Ibe et al. (2023), Mpagike and
Makungu (2024), Ataba et al. (2025). This is largely due to the
species’ ongoing domestication and the lack of structured development within
the aulacode (grasscutter) sector, coupled with the
limited scientific knowledge currently available Addo et al. (2007), Byanet et al.
(2009), Nteme et al.
(2009),(2010),(2014),
Onwuama et al.
(2018), Ibe et al. (2023), Mpagike and
Makungu (2024), Ataba et al.
(2025), Nteme et al.
(2025). Various methods of restraint and anesthesia
have been documented, including the use of acepromazine or ketamine alone or in
combination with xylazine Mensah
et al. (1992), Houben
et al. (2004), Mensah
et al. (2006), Abe et al. (2010), Vétérinaires
Sans Frontières. (n.d.).
As noted by other
authors Siter et
al. (1991), Mensah
et al. (1992), Annor et
al. (2009), Kingdon
et al. (2015) Vétérinaires
Sans Frontières. (n.d.), while one may question the impact of
captivity on animal behaviour (docility and aggressiveness), we consider this
impact to be minor or negligible compared with concerns related to feeding,
hygiene, and social interactions among individuals. In this context, it is
important to underscore that ethological and welfare considerations are
particularly critical for these rodents, as they are highly sensitive to
neglect in terms of nutrition and hygiene.
In the wild,
newborns in grasscutter follow their mother for at least one month. The most
common litter size in the grasscutter is four, although litters of eight or
more have also been reported Onadeko and Amubode (2002), Fantodji and
Soro (2004), Abioye
et al. (2008); Opara et
al. (2010). Birth weight ranges from 70 to 160 grammes,
and the sex ratio is generally balanced. Both the number and size of the
offspring appear to be influenced by the dam’s nutritional status Asibey et al.
(1974), Fantodji and
Soro (2004), with indications that larger females tend
to produce larger litters. The weight of each offspring, however, is largely
independent of its position within the uterine horn during embryonic
development Addo et al. (2003), Opara et
al. (2010).
Lactation is
crucial for the survival of newborns in mammals Hoshino
et al. (1979), Gittleman
and Thompson (1988), Hovey
and Trott (2004), Vallantin et
al. (2023). Within a few minutes after birth, the
mammary glands of the female grasscutter fill with milk, and the first suckling
occurs approximately thirty minutes after the birth of the last newborn. Young
grasscutter instinctively suckle and may feed from multiple mothers, which
ensures the survival of orphans. The frequency of suckling varies with age,
ranging from three to eight times per day at birth, to eight to fifteen times
per day shortly before weaning Fantodji and
Soro (2004).
Suckling gradually
decreases with age. Within three to four days after birth, the young begin
nibbling and consuming forage. During the fattening period, the juveniles
require less water, as they obtain most of their liquid from maternal milk or
the forage provided. In general, the young remain with their parents until
weaning, which occurs around one to one and a half months of age, after which
they are separated and adopt the adult diet Fantodji and
Soro (2004).
Overall, our
findings align with previous studies on the mammary apparatus of various
rodents, particularly the mouse and rat, which have been the most extensively
studied De et al. (2010), Ferrier
et al. (2012). As in all mammalian species Ferrier
et al. (2012), the mammary tissue of the grasscutter
develops along the ventral region of the trunk. Examination of the female
grasscutter’s mammary glands reveals both similarities and species-specific
differences compared to other rodents and lagomorphs. Notably, in the female
grasscutter, the mammary papillae (teats) are relatively short but noticeably
broader than those of commonly described domestic rodents. In addition, the
female grasscutter, like the female chinchilla, possesses three pairs of teats Carpentier
et al. (1994), Leck et al. (1998), Mayer et
al. (2004), Stekelorom-Parmentelat et al. (2006), which is fewer than in other species such
as the rabbit, mouse, and hamster, each of which has five pairs Crispens
et al. (1975), Tremblay
et al. (2002), Ferrier
et al. (2012), Rédaction et
al. (2017), and the rat, which has six pairs Tremblay
et al. (2001), De et al. (2010), Ferrier
et al. (2012). More specifically, as detailed in Table 1,
the adult mouse has three thoracic pairs, one abdominal pair, and one inguinal
pair, whereas the rat has one cervical pair, two thoracic pairs, two abdominal
pairs, and one inguinal pair Ferrier
et al. (2012). Additionally, in both rabbits and hamsters,
the number of teats can vary depending on the breed Barone
et al. (1973), Lebas et
al. (2002), Rédaction Mon
hamster. (2017).
Although both the
female grasscutter and the chinchilla possess three pairs of teats, their
anatomical arrangement differs. While both species have thoracic and abdominal
pairs, the female grasscutter has an additional pectoral pair, absent in the
female chinchilla (Table 1). These differences are significant and can be
explained by the intrinsic morphology and behaviour of the two species. For
instance, adult chinchillas are smaller than adult grasscutters. Chinchillas
can reach up to 25 cm in body length, with the tail adding approximately 15 cm,
and females are generally larger than males Hess et al. (2025). In contrast, the adult female grasscutter,
excluding tail length, averages 45 cm in body length and approximately 3 kg in
body weight Ettian et al.
(2009). Moreover, in chinchillas, only the anterior
pair of teats is considered functional, which poses a problem when the litter
contains more than two newborns Sciama et al.
(2001), Stekelorom-Parmentelat, M. D. (2006). This is not an issue in grasscutters, as
all teats are functional after parturition. Litter sizes vary in both species
(1–6 young in chinchillas; 1–14 young in grasscutters), but the most common
litter sizes are two in chinchillas and four to five in grasscutters Fantodji and Soro (2004), Houben
et al. (2004), Stekelorom-Parmentelat, M. D. (2006), Frohlich
et al. (2025). Furthermore, in the female grasscutter, the
mammary glands merge to form a broad layer of mammary tissue along the ventral
surface of the trunk (Figure 1 and Figure 2), representing a distinctive
feature compared with other rodents and lagomorphs. According to previous
authors Shimer
et al. (1903), Steyn et al. (2018),
Mpagike and
Makungu (2024), Nteme et al.
(2025), these characteristics likely reflect
anatomical adaptations related to the grasscutter’s herbivorous diet, habitat
(tall-grass humid savannas, floodplains, marshes, reed beds, and riverbanks),
locomotion (adaptation for running), and behaviour (absence of burrowing).
|
Table 1 |
|
Table 1 Topographical Differences in Teat Locations between the Female Grasscutter,
Female Chinchilla, Mouse, and Rat |
||||
|
Female |
||||
|
Pair by region |
Aulacode |
Chinchilla |
Mouse |
Rat |
|
Thryonomys swinderianus (Temminck, 1827) |
(Chinchilla
lanigera × Chinchilla brevicaudata) |
(Mus musculus) |
(Rattus rattus) |
|
|
Cervical |
0 |
0 |
0 |
1 |
|
Pectoral |
1 |
0 |
0 |
0 |
|
Thoracic |
1 |
1 |
3 |
2 |
|
Abdominal |
1 |
1 |
1 |
2 |
|
Inguinal |
0 |
1 |
1 |
1 |
|
Total |
3 |
3 |
5 |
6 |
As in other
species, such as the chinchilla Stekelorom-Parmentelat, M. D. (2006), the unique positioning of the teats in the
mother, together with the ventral orientation of the young’s mouths, determines
the specific nursing posture of the female grasscutter. While suckling, the dam
supports part of her weight on her forelimbs and hindlimbs to prevent excessive
flattening of the abdomen. This teat arrangement allows the young to nurse on
either side despite their ventrally positioned mouths. However, if the female
grasscutter lies on her flank, access to the lower row of teats becomes
extremely difficult or even impossible Ewer et al. (1969), Asibey et al.
(1974), Kingdon
et al. (1974), Opara et
al.(2010).
The anatomical
study of the mammary glands of the female grasscutter (Thryonomys swinderianus Temminck, 1827) highlights morphological
features that may, on the one hand, assist in species identification and, on
the other, enhance the understanding of the lactation process, while also
contributing to improved clinical veterinary practices in grasscutter farming.
Specifically, it has been observed that the young can nurse while their mother
is feeding or resting. However, the mammary glands can also be affected by
purulent conditions (mastitis) or ectoparasitic infestations, which are highly
detrimental to the lactating female and harmful to the development of the young
Fantodji and
Soro (2004). This is significant because certain
anatomical features have contributed to classifying the grasscutter within the
suborder Hystricomorpha (porcupine-like rodents) Wood et al. (1955), Weir et al. (1974), Roberts and Perry (1974), Addo et al. (2007), Granjon
et al. (2009), Yapi et al. (2013), alongside species such as the crested
porcupine (Hystrix cristata), the agouti (Dasyprocta aguti), and the guinea pig
(Cavia porcellus), which is also domesticated in sub-Saharan Africa Ngou et al. (1995), Mballa
et al. (2017). Membership in the suborder Hystricomorpha also distinguishes the grasscutter from the
rat (Rattus rattus), mouse (Mus musculus), gerbil (Gerbillinae),
and hamster (Cricetinae), which belong to the
suborder Myomorpha (rat-like rodents).
More importantly,
one of the key implications of this study concerns the possibility of
implementing cross-fostering strategies aimed at equalizing the size of litters
and/or adjusting the weight of young grasscutters in commercial farms. For
example, in pigs, this practice—where piglets are transferred from one mother
to another within the first few hours after birth—is common, especially in the
case of highly productive or hyperprolific sows.
Indeed, when there are more live-born piglets than available active teats, it
becomes necessary to optimize the management of litters. This is particularly
crucial when the average birth weight of the newborns is lower than the
generally observed average. Furthermore, it is worth noting that this practice
also continues in non-hyperprolific farms, as long as
they maintain good sanitary status. The goal here is to balance the number of
piglets in each litter, with minimal exchange between mothers Oliveras
et al. (2020), Vande et
al. (2021), Casanovas
and Gasa (2022), Lorente and Sanjoaquin
(2023), (2024).
Our results suggest that in cases of lactation difficulties or hyperprolific litters, the use of foster mothers should be
prioritized for female grasscutters.
Although this
study was conducted on five captive adult females—thereby minimizing biases
related to individual variability Nteme et al.
(2025)—the relatively small sample size may limit
the generalizability of the findings Ataba et al.
(2025). Additionally, these results should be
interpreted in light of histological findings, since in rodents, it has been
observed that within each mammary gland, the histology of the ducts and alveoli
varies according to the gland’s location, the animal’s age, the stage of its
oestrous cycle, and pregnancy Boorman
et al. (1990), Ferrier
et al. (2012). Furthermore, as sexual dimorphism in
grasscutters is minimal, preventing easy visual distinction between males and
females Van der Merwe, M. (2007), Houben
et al. (2004), Byanet et al.
(2009), Annor et
al. (2009), future studies should compare the mammary
apparatus across sexes. In species with limited sexual dimorphism, such as the
crested porcupine (Hystrix cristata), which has compound mammary glands forming
bilateral complexes and whose secretory products are conveyed via excretory
ducts to two latero-pectoral teats, male teats are notably smaller and shorter Lopez et
al. (2013), Mouzoun et al.
(2018).
Despite these
limitations, this study provides novel anatomical insights into the female
grasscutter’s mammary apparatus, enhancing understanding of its reproductive
biology. Together with known features of the female genital tract (Thryonomys swinderianus Temminck, 1827) Nteme et al.
(2025), these traits likely shape maternal
behaviour in captivity and should be considered to improve reproductive
management in this characteristically African rodent.
Conclusion
In general,
mammary glands can vary in number and in their positioning on the ventral
surface of the trunk from one female to another. Their conformation and
morphology may serve as distinguishing criteria between species. Furthermore,
it is generally accepted that the number of teats and mammary glands in a
female is proportional to litter size. The aim of this study was to
characterize the topography and morphometry of the mammary apparatus in the
adult female grasscutter. While our results confirm significant anatomical
similarities between the grasscutter and other animal groups, including
rodents, it should be emphasized that the mammary glands of the female
grasscutter exhibit specific anatomical features unique to this species. These
morphological peculiarities influence the lactation process and are
particularly important for species diagnosis, for improving clinical veterinary
practices in grasscutter farming, and for biomedical research. Collectively,
these findings provide insight into the behavioural and socio-feeding patterns
of this predominantly African rodent, whose captive breeding is expanding
across several sub-Saharan African countries. They also suggest implementing
cross-fostering strategies in grasscutter farming to manage litter sizes and
adjust the weight of young, particularly in cases of lactation difficulties or hyperprolific litters. Finally, they lay the foundation for
further research aimed at a more comprehensive understanding of grasscutter
anatomy and the development of comparative frameworks with related or relevant
species, including the lesser cane rat (Thryonomys gregorianus
Thomas, 1894), from which it must be distinguished; hunted species such as the
Central African wild rabbit (Poelagus marjorita) and the crested porcupine (Hystrix cristata);
and the guinea pig (Cavia porcellus).
ACKNOWLEDGMENTS
The authors express their sincere gratitude to:
The “Horizons francophones” Program – AUF/West Africa;
The collaborators at EISMV Dakar, UNA Abidjan, and ENVAA Nantes-Oniris;
The grasscutter breeders in Côte d’Ivoire and Benin;
All those who, directly or indirectly, contributed to the completion of this study.
REFERENCES
Abe, S. R. (2010). Parasites Rencontrés Chez L’aulacode (Thryonomys swinderianus) En Côte D’ivoire : Cas Du District D’abidjan [Thèse D’exercice En Médecine vétérinaire]. EISMV.
Abioye, F. O., Uda, A. C., Opara, M. N., Aju, P. C., and Onyema, M. C. (2008). Adaptability of Grasscutter (Thryonomys swinderianus) in Natural and Domestic Environments. In Forestry Association of Nigeria (Ed.), Proceedings of the Conference of Forestry Association of Nigeria, August 15–20 (537–550).
Addo, P. G., Awumbila, B., Awotwi, E., and Ankrah, N.-A. (2007). Comparative Characterization of the Oestrous Cycles of the Grasscutter (Thryonomys Swinderianus) and the Guinea pig (Cavia porcellus) by the Hystricomorph Vaginal Membrane Perforation Phenomenon. Livestock Research for Rural Development, 19.
Addo, P., Dodoo, A., Adjei, S., and Awumbila, B. (2003). Optimal Duration of Male-Female Exposure to Optimize Conception in the Grasscutter (Thryonomys swinderianus). Livestock Research for Rural Development, 15, 35–45.
Adjanohoun, E. (1992). Le Cycle Sexuel et la Reproduction de l’aulacode (Thryonomys Swinderianus Temminck, 1827). Mammalia, 56(1), 109–119.
Annor, S. Y., Adu, E. K., Donkor, J., Otsyina, H., and Ahiaba, J. (2009). Grasscutter production: A handbook. GTZ.
Annor, S., Ben, A., Aboagye, G. S., Boa-Amponsem, K., Djang-Fordjour, K. T., and Cassady, J. P. (2011). The Genetics of Morphological Traits in the Grasscutter. Livestock Research for Rural Development, 23. https://www.lrrd.org/lrrd23/8/Anno23167.htm
Annor, S., Iddisah, I., and Djang-Fordjour, K. T. (2013). Growth, Carcass and Behaviour Characteristics of Castrated and Intact male Grasscutters (Thryonomys swinderianus). Livestock Research for Rural Development, 25. https://www.lrrd.org/lrrd25/3/anno25050.htm
Asibey, E. O. A. (1974). The Grasscutter (Thryonomys Swinderianus Temminck). Symposia of the Zoological Society of London, 34, 161–170.
Ataba, A., Guintard, C., and Tchaou, M. (2025). Radiographic Study of the Anatomy and Preliminary Barium Gastrointestinal Transit in the Grasscutter (Thryonomys Swinderianus, Temminck 1827). BMC Veterinary Research, 21, 690. https://doi.org/10.1186/s12917-025-05149-1
Baptist, R., and Mensah, G. (1986). The Cane Rat—Farm Animal of the Future? World Animal Review, 60, 2–6.
Barone,
R. (1976).
Anatomie Comparée Des Mammifères Domestiques, Tome 4 : Splanchnologie.
Laboratoire d’anatomie ENV Lyon.
Barone, R., Pavaux, C., and Blin, P. C. (1973). Atlas D’anatomie Du Lapin. Masson.
Boorman, G. A., Eustis, S. L., Elwell, M. R., Montgomery, C. A. J., and MacKenzie, W. F. (Eds.). (1990). Pathology of the Fischer rat: Reference and Atlas. Academic Press.
Broalet, E., Tako, A., Zunon-Kipre, Y., Ouatara, D., and Kouakou, F. (2012). Sur l’anatomie de l’aulacode (Thryonomys swinderianus, Temminck, 1827): Revue de littérature. Morphologie, 96(314), 100. https://doi.org/10.1016/j.morpho.2012.08.080
Byanet, O., Onyeanusi, B. I., and Ibrahim, N. D. G. (2009). Sexual Dimorphism with Respect to the Macro-Morphometric Investigations of the Forebrain and Cerebellum of the Grasscutter (Thryonomys swinderianus). International Journal of Morphology, 27(2), 361–365. https://doi.org/10.4067/S0717-95022009000200010
Carpentier, F. (1994). Contribution à l’étude du Chinchilla Considéré Comme Animal de Compagnie [Thèse d’exercice en médecine vétérinaire]. ENV Lyon.
Casanovas, J., and Gasa, J. (2022). Le Protocole D’adoption, une Question Complexe à Normaliser. 3trois3.com.
Child, M. F. (2016). Thryonomys Swinderianus (Errata Version Published in 2017). The IUCN Red List of Threatened Species. https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T21847A22278009.en
Crispens, C. G. (1975). Handbook on the Laboratory Mouse. Thomas Publisher.
De Assis, S., Warri, A., Cruz, M. I., and Hilakivi-Clarke, L. (2010). Changes in Mammary Gland Morphology and Breast Cancer Risk in Rats. Journal of Visualized Experiments, 44, 2260. https://doi.org/10.3791/2260
Dyce,
K. M., Sack, W. O., and Wensing,
C. J. G. (1996). Textbook of veterinary anatomy (2nd ed.). Saunders.
Ettian,
M. K., Soulemane, O., and Tahoux,
T. M. (2009).
Influence du régime alimentaire sur l’intervalle de parturition des aulacodes
en captivité dans la région de Grand-Lahou (Côte d’Ivoire, Afrique de l’Ouest).
Journal of Animal and Plant Sciences, 4(1), 311–319.
Ewer, R. F. (1969). Form and function in the grasscutter Thryonomys swinderianus Temminck (Rodentia, Thryonomyidae). Ghana Journal of Science, 9, 131–149.
Fantodji, A., and Soro, D. (2004). L’élevage d’aulacodes : Expérience en Côte d’Ivoire. Éditions du GRET, MAE. https://gret.org/wp-content/uploads/2021/11/GP-19_Elevage-aulacodes.pdf
Ferrier, E. (2012). Étude du développement du tissu mammaire chez la souris adulte en région interscapulaire comparée à la région ventrale périmamélonnaire sous l’influence des œstrogènes [Thèse d’exercice en médecine vétérinaire, ENVA]. https://www.mcours.net/fra6/fatslfra6pan228.pdf
Frohlich, J. (2025). Chinchillas—Exotic and Laboratory Animals. MSD Veterinary Manual.
Gittleman, J. L., and Thompson, S. D. (1988). Energy Allocation in Mammalian Reproduction. American Zoologist, 28(3), 863–875. https://doi.org/10.1093/icb/28.3.863
Granjon, L., and Duplantier, J.-M. (with Quesseveur, E.). (2009). Les Rongeurs De l’Afrique Sahélo-Soudanienne. IRD. https://horizon.documentation.ird.fr/exl-doc/pleins_textes/ed-09-10/010048662.pdf
Grassé, P.-P.(Ed.). (1971). Traité de zoologie : Anatomie, Systématique, Biologie. Tome Xvi, Mammifères. Fascicule 3, Musculature des Membres, Musculature Peaucière, Musculature Des Monotrèmes, Arthrologie. Masson.
Hess, L. (2025). Description and Physical Characteristics of chinchillas—All other pets. MSD Veterinary Manual. https://www.msdvetmanual.com/all-other-pets/chinchillas/description-and-physical-characteristics-of-chinchillas
Hoshino, K. (1979). Hormonal Teratogenesis in Mammary Glands of the Mouse. In T. V. N. Persaud (Ed.), Teratological Testing (139–160). Springer Netherlands. https://doi.org/10.1007/978-94-011-6651-5_7
Houben, P. (1999). Elevage d’aulacodes au Gabon : Éléments de bilan. Canopée, 15 (octobre), 7–8.
Houben, P., Edderai, D., and Nzego, C. (2004). Élevage de l’aulacode : Manuel de l’éleveur (D. Cornélis, Ed.). CIRAD-EMVT.
Hovey, R. C., and Trott, J. F. (2004). Morphogenesis of Mammary Gland Development. Advances in Experimental Medicine and Biology, 554, 219–228. https://doi.org/10.1007/978-1-4757-4242-8_19
Ibe, C. S., Ogbonnaya, O., Ikpegbu, E., and Ani, N. V. (2023). Anatomical Studies on the African Grasscutter (Thryonomys swinderianus), a Key Component of the Minilivestock Industry in Nigeria. The Anatomical Record, 306(1), 226–234. https://doi.org/10.1002/ar.25049
Jori, F. (2001). La Cria De Roedores Tropicales (Thryonomys swinderianus y Atherurus africanus) Como Fuente De Alimento En Gabon, Africa Central [PhD thesis, Facultat de Veterinaria, Universitat Autònoma de Barcelona]. https://publications.cirad.fr/une_notice.php?dk=483517
Jori, F., and Cooper, J. E. (2001). Spontaneous Neoplasms in Captive African Cane Rats (Thryonomys swinderianus Temminck, 1827). Veterinary Pathology, 38(5), 556–558. https://doi.org/10.1354/vp.38-5-556
Jori, F., Cooper, J. E., and Casal, J. (2001). Postmortem Findings in Captive Cane Rats (Thryonomys swinderianus) in Gabon. Veterinary Record, 148(20), 624–628. https://doi.org/10.1136/vr.148.20.624
Kingdon, J. (1974). East African mammals: An Atlas of Evolution in Africa. 2. Part B, Hares and Rodents. Academic Press.
Kingdon, J. (2015). Kingdon Field Guide to African Mammals (2nd ed.). Bloomsbury Wildlife.
Kon, S. K., and Cowie, A. T. (Eds.). (1961). Milk: The Mammary Gland and its Secretion. Academic Press.
Lebas, F. (2002). Cuniculture : Biologie du lapin—Chapitre 2 : Extérieur du corps. Cuniculture.info.
Leck, S. (1998). Chinchillas: What Every Veterinarian Needs to Know. Exotic DVM, 1(1), 30–31.
Lopez, L. (2013). Atlas radiographique et Ostéologique Du Porc-épic : Hystrix indica [Thèse d’Exercice en Médecine Vétérinaire, ENVT].
Lorente, J., and Sanjoaquin, L. (2023). Adoptions et Nourrices (I): Faire face à l’hyperprolificité. 3trois3.com.
Lorente, J., and Sanjoaquin, L. (2024). Adoptions et Nourrices (II): Comment réaliser les Mouvements ? 3trois3.com.
Mayer, J. (2004). Natural History of the Chinchilla (Chinchilla lanigera). Exotic Mammal Medicine and Surgery, 2(1), 9.
Mballa, D. (2017). Afrique : L’élevage des Cobayes, un Juteux Business En Expansion. AfricTelegraph.
Mensah, G. A., Houinato, M., Koudanbe, O. D., Bembide, C., Dossou-Gbete, G. S. O., Mensah, S. E., Pomalegni, C. B., and Kpera, G. N. (2006). Fiche Technique : Castration De L’aulacode (Thryonomys swinderianus) mâle d’élevage. Bulletin de la Recherche Agronomique du Bénin, 54, 15–16.
Mensah, G. A., Stier, C. H., and Gall, C. F. (1992). Aspects Pratiques en élevage d’aulacodes (Thryonomys swinderianus). IV. Premiers Essais De Tranquillisants (per os). REMVT, 45(1), Article 1.
Mouzoun, S. (2018). Écologie et Connaissances Ethnozoologiques du Porc-Épic à crête (Hystrix cristata Linnaeus, 1758) Dans Les Réserves De Biosphère de la Pendjari et du W du Bénin [PhD thesis]. Université d’Abomey-Calavi.
Mpagike, F. H., and Makungu, M. (2024). Radiological Anatomy of the Pelvis and pelvic limb of the greater cane rat (Thryonomys swinderianus). Veterinary Medicine International, 2024(1), 5998717. https://doi.org/10.1155/2024/5998717
Ngou Ngoupayou, J. D., Kouonmenioc, J., Fotso Tagny, J. M., Cicogna, M., Castroville, C., Rigoni, M., and Hardouin, J. (1995). Possibilités de développement de l’élevage du cobaye en Afrique subsaharienne : Le cas du Cameroun. Revue Mondiale de Zootechnie, 83.
Nteme
Ella, G. S., Aoussi, A., Penda,
R., Fantodji, A., Missohou,
A., and Bakou, S. (2009). Composition chimique des muscles du Membre Pelvien Du Grand Aulacode
(Thryonomys Swinderianus, Temminck 1827). RASPA, 7(1), 29–32.
Nteme Ella, G. S., Aoussi, S., Bakou, S., Ouassat, M., and Costiou, P. (2010). Étude Descriptive Des Muscles De La Cuisse Du Grand Aulacode (Thryonomys swinderianus, Temminck 1827). Revue Africaine de Santé et de Productions Animales, 8(1), 41–46.
Nteme Ella, G. S., Aoussi, S., Kouame, P., Fantodji, A., and Bakou, S. (2014). Détermination de la couleur et de la valeur nutritionnelle de la viande d’aulacodes sauvages (Thryonomys swinderianus, Temminck, 1827). International Journal of Biological and Chemical Sciences, 8(2), 610–620. https://doi.org/10.4314/ijbcs.v8i2.19
Nteme
Ella, G. S., Kamga Waladjo,
A. R., Bakou, S., and Akakpo, A. J. (2005). Anatomical
Particularities of the Genital
Tract of the Female Grasscutter
(Thryonomys swinderianus).
Revue Africaine De Santé Et De Productions Animales, 3(2), Article 2.
Nteme Ella, G. S., Ntsama Abah, F., Aoussi, S., and Bakou, S. (2025). Particularités ostéologiques du pied du grand aulacode (Thryonomys swinderianus, Temminck, 1827). C@C, 13, 1–35.
Oliveras, A. (2020). Adoptions dans la pratique : Une étape clé pour maximiser la quantité et la qualité des sevrés. 3trois3.com.
Onadeko, S. A., and Amubode, F. O. (2002). Reproductive Indices and Performance of Captive Reared Grasscutters (Thryonomys swinderianus Temminck). Nigerian Journal of Animal Production, 29(1), 142–149. https://doi.org/10.51791/njap.v29i1.1548
Onwuama, K. T., Ojo, S. A., Hambolu, J. O., Dzenda, T., Zakari, F. O., and Salami, S. O. (2018). Macro-anatomical and Morphometric Studies of the Hindlimb of grasscutter (Thryonomys swinderianus, Temminck 1827). Anatomia, Histologia, Embryologia, 47(1), 21–27. https://doi.org/10.1111/ahe.12319
Opara, M. N. (2010). The grasscutter I: A Livestock of Tomorrow. Research Journal of Forestry, 4(3), 119–135. https://doi.org/10.17311/rjf.2010.119.135
Popesko,
P. (1972). Atlas
d’anatomie Topographique Des Animaux Domestiques: Vol.
I. Vander.
Raynaud, A. (1961). Morphogenesis of the mammary gland. In S. K. Kon and A. T. Cowie (Eds.), Milk: The mammary gland and its secretion (3–46). Academic Press.
Rédaction Mon hamster. (2017). Les premiers jours du hamster. Mon hamster.
Roberts,
C. M., and Perry, J. S. (1974). Hystricomorph embryology.
Symposium of the Zoological Society of London, 34,
333–360.
Sciama, Y. (2001). Le chinchilla. De Vecchi Editions.
Shimer, H. W. (1903). Adaptations to Aquatic, Arboreal, Fossorial and Cursorial Habits in Mammals. III. Fossorial Adaptations. The American Naturalist, 37(444), 819–825. https://doi.org/10.1086/278368
Siter, C.-H., Mensah, G. A., and Gall, C. F. (1991). Elevage d’aulacodes (Thryonomys swinderianus) pour la production de viande. Revue Mondiale de Zootechnie, 69(4). https://www.fao.org/4/u5700t/u5700t0j.htm
Soro, D. (2007). Stratégies de Conduite D’élevage Pour Des Performances de Reproduction Des Aulacodes D’élevage En Côte D’ivoire, Étude Intégrée de la physiologie sexuelle de l’aulacodin [PhD thesis, Université d’Abobo-Adjamé (UAA), UFR SN].
Stekelorom-Parmentelat, M. D. (2006). Le chinchilla, nouvel animal de compagnie [Thèse d’exercice en médecine vétérinaire, ENV Lyon]. https://www2.vetagro-sup.fr/bib/fondoc/th_sout/th_pdf/2006lyon010.pdf
Tondji, P. M., and Agbessi,
A. F. N. (1992). Données Générales
Sur L’anatomie de l’aulacode (Thryonomys swinderianus). In L’aulacodiculture:
Acquis et perspectives: Actes de la 1ère Conférence
Internationale, Cotonou (République du Bénin), 17–19 février 1992 (1, 49–71).
Tremblay,
M. (2001). Le
rat. Le Jour.
Tremblay, M. (2002). La Souris. Le Jour.
UCAD. (2024). Comité d’éthique de la recherche de l’UCAD. Portail de la Recherche – UCAD.
Vallantin, C. (2023). Aspects évolutionnistes et comparatistes de la lactation: Des Premiers Mammifères à nos jours (105) [Mémoire présenté pour l’obtention du Diplôme d’État de Sage-Femme]. École de Maïeutique de Marseille.
Van der Merwe, M. (2007). Discriminating between Thryonomys Swinderianus and Thryonomys Gregorianus. African Zoology, 42(2), 165–171. https://doi.org/10.10520/EJC18054
Vande Pol, K. D., Bautista, R. O., Harper, H., Shull, C. M., Brown, C. B., and Ellis, M. (2021). Effect of Within-Litter Birth Weight Variation After Cross-Fostering on Piglet Preweaning Growth and Mortality. Translational Animal Science, 5(3), txab039. https://doi.org/10.1093/tas/txab039
Vétérinaires Sans Frontières. (n.d.). Formation en aulacodiculture: La prévention des maladies—Les soins. Projet DGEG. Retrieved 12 July 2025
Weir, B. J. (1974). Reproductive Characteristics of Hystricomorph Rodents. Symposium of the Zoological Society of London, 34, 265–301.
Wood, A. E. (1955). A Revised Classification of the Rodents. Journal of Mammalogy, 36(2), 165–187. https://doi.org/10.2307/1375874
Yapi, Y. M. (2013). Physiologie digestive de l’aulacode (Thryonomys swinderianus) en croissance et impact des teneurs en fibres et céréales de la ration sur la santé et les performances zootechniques [Doctorat, Institut National Polytechnique de Toulouse (INPT)]. HAL. https://theses.hal.science/tel-04314022v1/file/yapi.pdf
Yewadan, L. T. (1992). Feeding of aulacodes (Thryonomys swinderianus, Temminck 1827) Reared in Narrow Captivity. In L’aulacodiculture: Acquis et perspectives: Actes de la 1ère Conférence Internationale, Cotonou (République du Bénin), 17–19 février 1992 (1, 143–149).
 This work is licensed under a: Creative Commons Attribution 4.0 International License
This work is licensed under a: Creative Commons Attribution 4.0 International License
© Granthaalayah 2014-2025. All Rights Reserved.