ALLELOPATHIC EFFECT OF THREE WEED SPECIES ON THE GERMINATION AND SEEDLING GROWTH OF TOMATO (LYCOPERSICON ESCULENTUM)Adeleke, Martina T. V 1 1,2 Department of Plant Science and Biotechnology, Rivers State University, Nkpolu, Port Harcourt, Nigeria |
|
||
|
|
|||
|
Received 05 January 2022 Accepted 11 February 2022 Published 28 February 2022 Corresponding Author Adeleke, Martina T. V, mtadeleke@yahoo.co.uk DOI 10.29121/granthaalayah.v10.i2.2022.4490 Funding: This research
received no specific grant from any funding agency in the public, commercial,
or not-for-profit sectors. Copyright: © 2022 The
Author(s). This is an open access article distributed under the terms of the
Creative Commons Attribution License, which permits unrestricted use, distribution,
and reproduction in any medium, provided the original author and source are
credited.
|
ABSTRACT |
|
|
|
Laboratory and green house experiments
were conducted to ascertain the effect of exudates (from the vegetative parts
of the weeds soaked in water) of different ages (36hrs, 72hrs and 120hrs)
from three common weed species (namely, Chromolaena odorata, Ageratum conyzoides and Cyperus esculentus) on the seed
germination and seedling growth of tomato (Lycopersicum esculentum). Tomato seeds were sown
in petri-dishes on filter paper moistened with the treatments, and another
set of tomato seeds were sown in soil, and treatments applied. The exudates
of C. odorata had the greatest
inhibitory effect on seed germination on filter paper, while those of A. conyzoides
followed closely. C. esculentus had
the least inhibitory effect. The weed exudates however did not have marked
inhibition on seed germination and seedling growth in the soil medium;
rather, they competed favorably with the Control. It was observed that the
longer the soaking duration of the weeds in water, the less inhibitory their
exudates were on seed germination, with water dilution. All the three weed
species considered showed allelopathic effects, but especially C. odorata and A. conyzoides when in direct contact
with tomato seeds. |
|
||
|
Keywords: Weeds, Exudate, Chromolaena Odorata,
Ageratum Conyzoides, Cyperus Esculentus, Tomato 1. INTRODUCTION
Weed management has been a challenge, and its management with the use
of herbicides is fast becoming least desired because of the pressure from
society for healthier farm produce Kong (2010). There is therefore the need for other means of
weed management such as cultural, physical, biological, mechanical, and maybe
a combination of all these to avoid the use of herbicides Rizzardi (2011). Weeds are
also known to have allelopathic effects as part of their competition with
other plants. Allelopathy, according to Inderjit and Callaway (2003) has to do with effects a plant could have (directly
or indirectly) on other plants around it, which could either be beneficial or
harmful. A plant is capable of this by releasing chemicals it produces into
the surrounding environment, by its root exudates, leaching, decomposition or
volatilization. Put in other words, allelopathy could be said to be a natural
biological defense mechanism of plants for their growth, reproduction, and
survival, which they try to achieve by producing some biochemicals Roger et al. (2006). Furthermore, allelopathy, which is intrinsic in
some plants, stimulates or stops the survival of neighboring plants by this
ability to produce allelochemicals Javed (2020). This ability of these chemical compounds to hinder
the growth of other plants depends on its |
|
||
concentration in the
allelopathic plants, and how susceptible those around it are Koocheki et al. (2001). This phenomenon of
allelopathy has been growing in interest, especially now, in relation to
solving the weed problem Rice (1984), Marcı et al. (2004), Vasilakoglou et al. (2005), Dhima et al. (2006).
As a natural phenomenon,
allelopathy is a welcome idea that could be used in weed and pest management in
cultivated fields.
Among the water soluble
allelochemicals are derivatives of phenols, flavonoids, coumarins, alkaloids,
terpenoids, ethylene’s and quite a number of other secondary metabolites which
can stop or stimulate the germination and growth of plants and other organisms
in the environment Rashid
et al. (2010), Fujji et al. (2004). The deleterious
action of these allelochemicals could be in different ways such as on the
physiology, enzymatic activities, germination and even synthesis Vasconcelos
et al. (2012). Hence the serious
consideration of allelopathy as a tool in managing some weed species in
farmlands. Rezende
et al. (2003), Kong (2010).
Chromoleana odorata (Siam weed) and Ageratum conyzoides
(Billy goat weed), both of the family Asteraceae; and Cyperus esculentus (Yellow nutsedge) of
the family Cyperaceae, are common weeds of the
tropics, growing aggressively in cultivated fields. C. odorata is a diffuse, rapidly growing, strongly scented
perennial shrub, which reproduces from seeds and vegetative parts. Its
invasiveness and aggressive growth can be tied not just to its heavy seed
production, but also to the large amounts of allelochemicals such as flavonoids
and phenols in its leaves. These are known to affect seedling germination Zachariades et al. (2009), Eze and Gill (1992).
A. conyzoides
is an erect, hairy aromatic annual herb that reproduces from seeds, while C. esculentus, is a tall, rhizomatous,
tuber-bearing perennial sedge which reproduces mainly vegetatively from
nutlets, and sometimes by seeds that germinate while on the spikes.
Tomatoes, on the other hand, is considered one of the fast-germinating plants
that is sensitive to secondary metabolites in the environment, so it can be
used as a bioindicator of weed allelopathic activity Ferreira
and Áquila (2000), Cândido et al. (2010).
The aim of this study is
therefore to identify possible allelopathic effects of three different strengths
of exudates from the weeds: Cyperus
esculentus Lin. (Yellow nutsedge), Chromolaena odorata
L. (Siam weed) and Ageratum conyzoides Linn (Billy goat weed), on seed germination
and seedling growth of Lycopersicum esculentum L. (tomato).
2. MATERIALS AND METHODS
Laboratory and green house experiments were conducted
at the Department of Plant Science and Biotechnology, Rivers State University,
Port Harcourt, Nigeria. For the production of the
exudates, the weeds: Cyperus esculentus,
Chromoleana odorata and Ageratum conyzoides were collected by
uprooting in the Teaching and Research Farm of the Rivers State University.
Tomato fruits were purchased from the market, and their seeds separated, washed,
and air-dried.
2.1. PREPARATION OF WEED EXUDATE
The collected weeds were washed free of soil and
air-dried. One kilogram of each weed type was weighed out on a weighing scale,
and the whole plant- leaves, stem, and roots, were sliced into small 2cm pieces
with a kitchen knife, and then soaked in 1 litre of distilled water. This was
done in three replicates for each weed in plastic bowls: giving a total of nine
bowls, well labelled. Each weed was soaked for varying periods of time viz: 36,
72 and 120 hours. These different durations of soaking the weed species
constitute the treatments which were administered on the tomato seeds.
After 36 hours of soaking, the
weed exudate was sieved using a cloth sieve and the extract stored in a
refrigerator until used. The same thing was done with the second set of soaked
weeds after 72 hours, and with the third set after 120 hours.
2.2. LABORATORY EXPERIMENT
Sets of 20 tomato seeds were each
arranged on Whatman No. 1 filter paper placed in petri dishes. Nine petri
dishes were arranged per treatment, and a set of control, making it 30
petri-dishes in all. The weed exudates were retrieved from the refrigerator and
allowed to come to room temperature. They were then administered on the filter
paper (with seeds on them); 4ml each of treatment exudate solution was applied.
Only distilled water was used for the control.
The petri dishes were place near the window at room temperature (28 ± 2oC).
Germination counts were made with emergence of 1mm of radicle. The filter
papers were constantly moistened with the appropriate exudate until the eighth
day after commencement of the experiment; subsequently, 2ml of distilled water
was used to moisten the filter paper. Germination counts and measurement of the
radicle of germinating seedlings continued until the fifteenth day.
2.3. GREENHOUSE EXPERIMENT
Polyethylene bags of 1kg capacity
were half-filled with 500g of air-dried loamy soil. Three replicates for each
of the three weed exudates for three treatments (36, 72 and 120 hours) were
prepared, plus a set of three replicates for the control, making a total of 30
bags. The tomato seeds were sown in the bags, and 50ml of the different
exudates were added, while distilled water was used for the control.
Subsequently, the bags were watered every other day with 100ml of distilled
water.
Germination counts were made, and plant height measurement of the seedlings
were taken.
2.4. DESIGN OF THE EXPERIMENT
The experiment was conducted in a randomized complete block design with three weed species (C. odorata, A. conyzoides and C. esculentus) and three treatments (weeds soaked for 36, 72 and 120 hours in distilled water), replicated three times each. Two experiments were conducted with three species and three treatments in a 3x 3 form and a control: one in the petri- dishes and the other in poly bags (a greenhouse experiment).
3. RESULT AND DISCUSSION
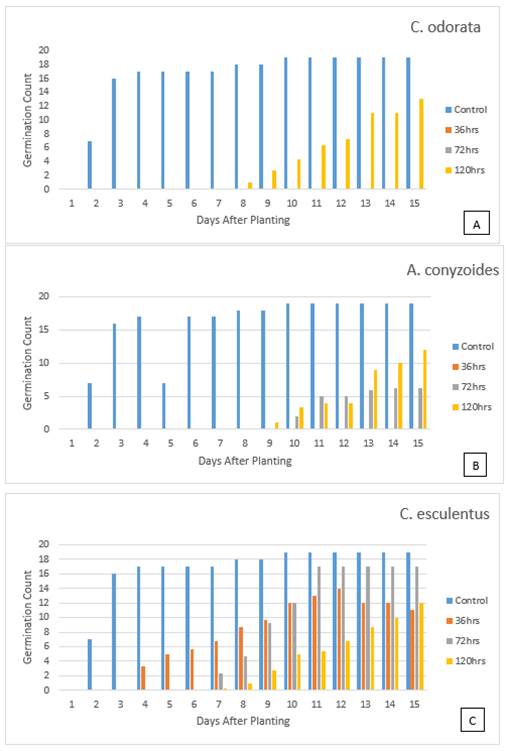
The effect of the weed exudates was much more
pronounced in the petri dish germination experiment than in that of soil.
Furthermore, the weed exudates, especially of C. odorata and A. conyzoides, soaked in distilled water for just 36hrs had
more adverse effect on germination of the tomato seeds; and the adverse effect
eased out, the older the exudate, as seen by the increase in germination counts
Table 1, Figure 1 and Figure 2. Even then, there was
no germination until the seventh day, just before the treatment concentrations
were diluted with the addition of 2ml of distilled water to moisten the filter
paper, then germination commenced in earnest. However, the seeds in the 36
hours exudate of A. conyzoides,
and 36 and 72 hours of C. odorata
still didn’t grow at all, even when diluted (Figure 1 & Figure 2). Leachates from
plants have been shown to suppress seed germination, vegetative propagules, and
early seedling growth Babu and Kandasanmy (1997), Dhwan and Gupta (1996), and decrease radicle
growth Casado (1995).
Of the three weed species used, C. esculentus had the mildest adverse effect on germination; even
with the 36 hours old exudate, it competed favourably with the control in
germination count: it started on the fourth day, while the control started on
the second day Table 1, Figure 1).
This general inhibition of seed germination by the treatments until water
dilution, points to the fact that these weed exudates all have allelochemicals
in them which inhibits tomato seed germination; C. odorata having the strongest inhibition. Eze and Gill (1992) reported that C. odorata contains a large amount of
allelochemicals, especially in the leaves, which inhibits the growth of many
plants in nurseries and plantations.
|
Table 1 Treatment means of the Germination
Counts (in petri dishes) of the three weed exudates |
||||||||||
|
|
|
ExudateA |
ExudateB |
ExudateC |
||||||
|
DAP |
Control |
Trmt1 |
Trmt2 |
Trmt 3 |
Trmt |
Trmt2 |
Trmt 3 |
Trmt |
Trmt2 |
Trmt3 |
|
1 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
2 |
7 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
3 |
16 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
4 |
17 |
- |
- |
- |
- |
- |
- |
3.3 |
- |
- |
|
5 |
17 |
-
|
- |
- |
- |
- |
- |
5 |
- |
- |
|
6 |
17 |
- |
- |
- |
- |
- |
- |
5.7 |
- |
- |
|
7 |
17 |
- |
- |
- |
- |
- |
- |
6.7 |
2.3 |
0.3 |
|
8 |
18 |
- |
- |
1 |
- |
- |
- |
8.7 |
4.7 |
1 |
|
9 |
18 |
- |
- |
2.7 |
- |
- |
1 |
9.7 |
9.3 |
2.7 |
|
10 |
19 |
- |
- |
4.3 |
- |
2 |
3.3 |
12 |
12 |
5 |
|
11 |
19 |
- |
- |
6.3 |
- |
5 |
4 |
13 |
17 |
5.3 |
|
12 |
19 |
- |
- |
7.3 |
- |
5 |
4 |
14 |
17 |
6.7 |
|
13 |
19 |
- |
- |
11 |
- |
6 |
9 |
12 |
17 |
8.7 |
|
14 |
19 |
- |
- |
11 |
- |
6.3 |
10 |
12 |
17 |
10 |
|
15 |
19 |
- |
- |
13 |
- |
6.3 |
12 |
11 |
17 |
12 |
|
|
|
Figure 1 The effect, with age, of
exudate from different weed species on germination of tomato seeds |
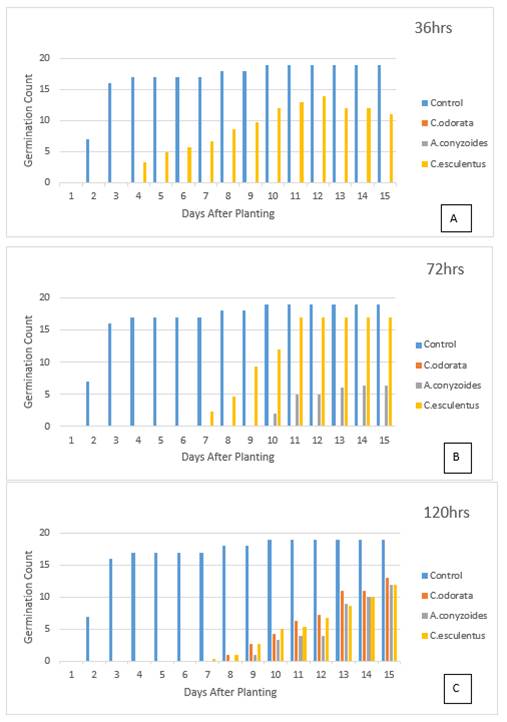
A. conyzoides which was next to C. odorata in inhibition, allowed the germination of some tomato seeds with dilution of the 72- and 120-hour exudate, but not 36 hr. This similar effect it had as C. odorata is expected since they both belong to the same family, Asteraceae. Members of the Asteraceae family have aromatic polyacetylene compounds. Polyacetylene compounds of this family have cytotoxic, antimicrobial, anti-inflammatory, neurotoxic, phototoxic, and several other types of activity Konovalov (2015). Natural polyacetylenes are compounds whose structures contain two or more triple bonds Christensen (1998). In the wider sense, acetylene compounds include all substances with carbon-carbon triple bonds or alkynyl functional groups. Compounds containing triple bonds are relatively unstable compounds like other unsaturated organic substances; and they are chemically and biologically active. The high reactivity leads to rapid oxidation and degradation of these compounds, especially on exposure to UV light. Konovalov (2015). This probably explains the observation in Table 1 that the longer the weeds were soaked (120 hrs), the lower the inhibitory effect of the exudate with dilution. Hence seeds of C. odorata germinated with dilution only at 120 hours and not before; and the same was observed with the radicle length in Table 2. With exposure therefore, the polyacetylene compounds were oxidized and degraded from the plant residue, thus reducing its inhibitory effect Gill et al. (1993).
C. esculentus on the other
hand, is a member of family Cyperaceae commonly known
as sedges. In the last few decades, constituents with great chemical diversity
were isolated from sedges, and a wide range of biological activities were
detected either for crude extracts or for pure compounds. Among the isolated
compounds, phenolic derivatives are the most important, especially stilbenoids, and flavonoids. To date, more than 60 stilbenoids were isolated from 28 Cyperaceae
species. Pharmacological investigation of Cyperaceae stilbenoids revealed that several compounds possess
promising activities: mainly antiproliferative, antibacterial, antioxidant and
anthelmintic effects. Moreover, stilbenes are important from chemotaxonomical point of view, and they play a key role in
plant defence mechanisms as well Dávid et al.
(2021).
|
|
|
Figure 2 The effect of ageing weed
exudates on the germination count of tomato seeds |
Germination count was highest in the 72 hrs exudate of C. esculentus (Figure 1C), but highest in the 120hrs exudate for C. odorata and A. conyzoides (Figure 1 A and Figure 1B). The germination count for all three weed treatments were however almost the same at 14 and 15 days after planting (Figure 2 C) in 120 hrs exudate, but that of C. odorata was slightly higher.
Root length measurement of germinating seedlings in C. esculentus showed increase in length with increase in the duration of soaking the weed Table 3, but with dilution. Again, this could be as a result of the decomposition of the plant residue which provides more nutrients for seedling growth with dilution.
|
Table 2 Effect of weed exudate treatments on radicle length of tomato (mm) |
|||||
|
TRMTS |
DAP |
Control |
C. odorata |
A. conyzoides |
C. esculentus |
|
36hrs |
Day
3 |
10.4 |
- |
- |
- |
|
Day
7 |
15.2 |
- |
- |
4.17 |
|
|
Day
11 |
18.2 |
- |
- |
8.48 |
|
|
Day
15 |
31.4 |
- |
- |
9.2 |
|
|
72hrs |
Day
3 |
12.4 |
- |
- |
- |
|
Day
7 |
26.6 |
- |
- |
- |
|
|
Day
11 |
41.1 |
- |
- |
17.7 |
|
|
Day
15 |
49.3 |
- |
- |
20.5 |
|
|
120hrs |
Day
3 |
17.5 |
- |
- |
- |
|
Day
7 |
27.2 |
10.2 |
17 |
24.4 |
|
|
Day
11 |
42.2 |
18.5 |
24 |
27.6 |
|
|
Day
15 |
46.3 |
22.6 |
23 |
28.8 |
|
Greenhouse Experiment
The percentage germination of
the tomato seeds in soil reveals that the treatment effect of the various
exudates is not as pronounced as that in the petri dish. This could be as a result of the interactions of the various soil
parameters, such as: soil temperature, soil moisture regime, alternate wetting
and drying of soil, soil nitrate level, among others, that affect seed
germination Akobundu (1987).
|
Table 3 Treatment Effect on the Germination percentage of Tomato seeds sown in soil |
|||||
|
DAP |
Control |
C. odorata |
A. conyzoides |
C. esculentus |
|
|
36hrs |
5 |
35 |
30 |
30 |
40 |
|
10 |
40 |
35 |
35 |
40 |
|
|
15 |
45 |
45 |
50 |
50 |
|
|
72hrs |
5 |
35 |
40 |
30 |
30 |
|
10 |
40 |
40 |
40 |
40 |
|
|
15 |
45 |
45 |
45 |
45 |
|
|
120hrs |
5 |
35 |
30 |
30 |
40 |
|
10 |
40 |
40 |
40 |
40 |
|
|
15 |
45 |
45 |
40 |
40 |
|
|
Table 4 Treatment effect on plant height (mm) of tomato seedlings sown in soil 15DAP |
||||
|
Trmts |
Control |
C. odorata |
A. conyzoides |
C. esculentus |
|
36hrs |
86.3 |
106 |
96.7 |
94 |
|
72hrs |
86.3 |
103 |
87.8 |
88.7 |
|
120hrs |
86.3 |
93.8 |
91.5 |
92.2 |
The plant height of the seedlings likewise reveal that
all the treatments competed favourably with the control Table 4. This means that the
weed exudates had no inhibitory effect on seedling growth in the soil medium.
This could be because the roots do not come in close contact with the exudate
as much as they do on filter paper in the petri dish; as, according to Patrick
(1971) the extent of damage
to the crop is related to the degree of contact of roots to the exudates. In soil,
the exudate, after application would naturally drain down through the soil particles,
and more so with further wetting.
In conclusion, the exudates from C. odorata had the most adverse effect on tomato seed germination on filter paper in petri-dish, followed closely by A. conyzoides of the same family Asteraceae. C. esculentus had the least adverse effect on tomato seed germination of the three weed exudates used. The tomato seeds sown in soil didn’t have any significant difference in germination percentage and growth from the control. Turnover of these weeds therefore into fields being prepared for crop cultivation should be avoided, except they are left for extended period of time for oxidation and degradation of polyacetylene and phenolic compounds, as close contact of the fresh exudates of these weeds with seeds could delay or completely prevent germination. On the other hand, the polyacetylene compounds in C. odorata and A. conyzoides, though relatively unstable, if extracted could serve as a natural herbicide.
REFERENCES
Akobundu, J. O. (1987) Weed science in the topics : Principles and practices. John Whiley and sons : pp 522
Babu, C. M., Kandasanmy, O. S., (1997). Allelopathic effects of Eucalyptus globulus Labill.on Cyperus rotundus L. and Cynodon dactylon L. pers. Journal of Agronomy and crop science 179,2, 123 - 126. Retrieved from https://doi.org/10.1111/j.1439-037X.1997.tb00507.x
Casado, C. M., (1995). Allelopathic effect of Lantana camara (Verbenaceae) on Morning glory (Ipomoea tricolor) Rhodora, 97 :891, 264 - 274. Retrieved from https://www.jstor.org/stable/23313216
Christensen, L. P. (1998) Rec. Res. Dev. Phytochemistry, 2, 227 - 257.
Cândido ACS et al. (2010). Potencial alelopático de lixiviados das folhas de plantas invasoras pelo método sanduiche. Revista Brasileira de Biociências 8 : 268-272. Retrieved from https://www.seer.ufrgs.br/rbrasbioci/article/view/114940
Dhima KV, Vasilakoglou IB, Eleftherohorinos IG, Lithourgidis AS (2006). Allelopathic potential of winter cereals and their cover crop mulch effect on grass weed suppression and corn development. Crop Science 46 :345-352. Retrieved from https://doi.org/10.2135/cropsci2005-0186
Dhwan, S. R. Gupta, S. K. (1996). Allelopathic potential of various leachate combinations towards SG and ESG of Parthenium hysterophorus Linn. World - Weeds 3 :1, 85 -88.
Dávid, C. Z., Hohmann, J., & Vasas, A. (2021). Chemistry and Pharmacology of Cyperaceae Stilbenoids : A Review. Molecules (Basel, Switzerland), 26(9), 2794. Retrieved from https://doi.org/10.3390/molecules26092794
Eze JMO, Gill LS (1992). Chromolaena odorata a problematic weed. Compositae Newsletter 20 :14-18.
Ferreira AG, Áquila MEA. (2000). Alelopatia : uma área emergente da ecofisiologia. Revista Brasileira de Fisiologia Vegetal 12 : 175-204.
Fujji Y et al. (2004). Assessment method for allelopathic effect from leaf litter leachates. Weed Biology and Management 4 : 19-23. Retrieved from https://doi.org/10.1111/j.1445-6664.2003.00113.x
Gill LS, Anoliefo GO, Iduoze UV (1993). Allelopathic effect of aqueous extracts of siam weed on growth of cowpea. Chromoleena Newsletters 8.1.
Inderjit, Callaway RM. (2003). Expérimental design for the study of allelopathy. Plant and Soil 256 : 1-11. Retrieved from https://doi.org/10.1023/A:1026242418333
Javed K (2020). Allelopathy : A brief review. Journal of Novel Applied Sciences 9(1) : 1-12. Retrieved from https://www.researchgate.net/publication/338854230_ALLELOPATHY_A_BRIEF_REVIEW
Kong CH. (2010). Ecological pest management and control by using allelopathic weeds (Ageratum conyzoides, Ambrosia trifida, and Lantana camara) and their allelochemicals in China. Weed Biology and Management,10 : 73-80. Retrieved from https://doi.org/10.1111/j.1445-6664.2010.00373.x
Konovalov, D. A. (2015) Polyacetylene Compounds of Plants of the Asteraceae Family (Review). Pharmaceutical Chemistry Journal 48(9) : 36-53 Retrieved from https://doi.org/10.1007/s11094-014-1159-7
Koocheki A, Zarif Ketabi H, Nakhforoush A (2001). Ecological approaches to weeds management. (Trans.), Ferdowsi University of Mashad Press, Iran.
Marcı'as FA Galindo JLG, Molindo JMG, Cutler HG (ed.) (2004). Allelopathy: Chemistry and mode of action of allelochemicals. CRC Press, New York.
Patrick, Z. A., (1971). Phytotoxic substances associated with the decomposition in soil of plant residues. Soil Sc. III : 13 - 18. Retrieved from https://doi.org/10.1097/00010694-197101000-00002
Rashid MH et al. (2010). The allelopathic potential of kudzu (Pueraria montana). Weed Science,58 : 47-55. Retrieved from https://doi.org/10.1614/WS-09-106.1
Rezende CP et al. (2003). Alelopatia e suas interações na formação e manejo de pastagens. Boletim Agropecuário. Lavras : UFLA. p. 1-55.
Rice EL (1984). Allelopathy. Academy Press: New York, NY. 1-6. Retrieved from https://doi.org/10.1016/B978-0-08-092539-4.50007-1
Rizzardi MA. (2011). Associação de herbicidas no manejo de plantas daninhas resistentes. Revista Plantio Direto 122 : 117-127.
Roger, M. R., Reigosa, J., Pedrol, N. and Gonzalez L. (2006) Allelopathy: A Physiological Process with Ecological Implications. Springer, pp 1, ISBN 1402042795 Retrieved from https://doi.org/10.1007/1-4020-4280-9
Vasconcelos MCC et al. (2012). Interferência de plantas daninhas sobre plantas cultivadas. Revista Agropecuária Científica no Semiárido 8 : 1-06.
Vasilakoglou I Dhima K, Eleftherohorinos I (2005). Allelopathic potential of Bermuda grass and johnson grass and their interference with cotton and corn. Agronomy Journal 97 :303 313 Retrieved from https://acsess.onlinelibrary.wiley.com/doi/abs/10.2134/agronj2005.0303a
Zachariades C, Day M, Muniappan R, Reddy GVP (2009). Chromolaena odorata (L.) King and Robinson (Asteraceae). In Biological Control of Tropical Weeds Using Arthropods (eds. Muniappan R, Reddy GVP, Raman A), pp. 130-162, Cambridge University Press, Cambridge, UK Retrieved from https://doi.org/10.1017/CBO9780511576348.008
 This work is licensed under a: Creative Commons Attribution 4.0 International License
This work is licensed under a: Creative Commons Attribution 4.0 International License
© Granthaalayah 2014-2022. All Rights Reserved.