
|
|
|
|
IN VITRO PROPAGATION OF VACHELLIA MACRACANTHA, AN IMPORTANT SPECIES OF THE SEASONALLY DRY TROPICAL FOREST IN NORTHERN PERUSosa-Amay
Rosa M. 1, Delgado-Paredes Guillermo E. *1, 2
|
|
|
|


Article Type: Research Article
Article Citation: Sosa-Amay Rosa M, Delgado-Paredes Guillermo E, Vásquez-Díaz
Cecilia, Zuñe-Da Silva Felipe, and Rojas-Idrogo Consuelo. (2020). IN VITRO PROPAGATION OF VACHELLIA
MACRACANTHA, AN IMPORTANT SPECIES OF THE SEASONALLY DRY TROPICAL FOREST IN
NORTHERN PERU. International Journal of Research -GRANTHAALAYAH, 8(11), 371-380.
https://doi.org/10.29121/granthaalayah.v8.i11.2020.2502
Received Date: 15 November 2020
Accepted Date: 30 November 2020
Keywords:
Auxins-Cytokinins Interaction
Callus Induction
Clonal
Propagation
Nodal Segments
ABSTRACT
Vachellia macracantha (ex Acacia macracantha) is one of the most important tree species in the Seasonally Dry Tropical Forest (SDTR) of northern Peru. The study aims to develop an efficient callus induction and micropropagation protocol from microcuttings of adult trees of this mentioned species, distributed across the North of Peru to the South of Ecuador. Shoot and uninodal microcuttings from three to five-year-old plants were investigated respecting their vegetative propagation and rooting capacity in vitro. High levels of NAA (2.5 to 7.5 mg L-1) were most efficient for shoot elongation and axillary bud development, although the basal callus formation and defolitation occurrence was also very high. In this case, the NAA 0.02 mg L-1, BAP 0.05, and GA3 0.05 mg L-1 interaction has been produced a better multiplication rate after 30 days without both basal callus formation and defoliation. Our findings suggest that other treatments tested such as NAA-BAP, NAA-KIN and NAA-2iP interaction, not were significantly satisfactory. Rooting response and consequently shoot elongation were always better with IBA (0.5 to 2.0 mg L-1) than NAA, under the same growth regulators concentrations.
Abbreviations: BAP, 6-benzylaminopurine; NAA, α-naphthaleneacetic acid; IBA, 4-[3-indolyn] butyric acid; GA3, gibberellic acid; KIN, kinetin; 2iP, N6-[2-Isopentenyl] adenine.
1. INTRODUCTION
The genus Acacia belongs to the family Fabacaeae (Leguminosae), compring more than 1.350 species, distributed throughout tropical and warm temperate areas around the world [1]. Vachellia macracantha (Humb. & Bonpl. ex Willd.) Seigler & Ebinger (ex Acacia macracantha) is a forest tree species well adapted to water stress conditions and like most species of the genus Acacia have the natural ability to fix atmospheric nitrogen [2]. It grows in the seasonally dry tropical forest of northern coast of Peru, especially at the Pampas de Olmos, Lambayeque and Piura (Lat. 3º and 8º South) with precipitation between 100 and 600 mm and average temperature of 20 to 25 oC, on flat or rolling terrain, sandy or rocky soil generally derived from eolian or alluvial material from mountain streams, pH 7 to 8.2. V. macracantha, is populary named by locals as “faique”, “espino” or “huarango” [3], and in various pioneering reports it have been considered of greatest importance to reforest dry wastelands, erosion control and stabilization of coastal sand dunes, due to their salt tolerance and nitrogen fixation capacity [4]. Its fruits and leaves are a source of animal fodder and the stems are utilized as fuelwood, timber, among other benefits for humans and animals.
The existence of very heterogeneous stands as a consequence of the use of seeds in artificial plantations seems to be one of the most serious limitations in terms of fruit and woody productivity. In addition, the multiplication of germplasm, selections and massive propagation systems are therefore necessary to attain high numbers of desirable clones [5]. This problem is compounded due to self-incompatibility/outcrossing pollination inherent within several genus of leguminous plants, resulting in numerous interspecific hybrids found in natural populations[6].
Micropropagation, which is often used successfully for the multiplication of herbaceous and woody plants, represents an interesting alternative for this species, while among woody plants, regeneration from adults’ subjects is often not successful directly using this method [7]. However, in the last 30 years, the vegetative multiplication of woody species, has been rarely obtained from mature trees of several species of Vachellia (Acacia) such as in A. albida [8]and in gum arabic tree (A. senegal), which finfings show that nodal segments of plants produced during four years in a greenhouse induced the highest multiplication rate in MS culture medium with 5.0x10-5 M zeatin or BAP[9].
Most studies regarding micropropagation of Acacia species have been started from nodal segments of seedlings of various ages. In A. albida, cotyledonary nodes obtained from seedlings induced multiple shoots in MS culture medium supplemented with 0.5 mg L-1 NAA and 3.0 mg L-1 BAP [10]. In A. auriculiformis, a native species of Papua New Guinea and northern Australia, the in vitro micropropagation was observed in nodal segments from 5-months-old seedlings in MS culture medium supplemented with 0.44 μM (0.5 mg L-1) BA, yielding 10 shoots/explant, and the investigations also recorded that MS medium produced a higher number of shoots compared to both WPM [11] and B5[12] culture medium [13]. Despite that these authors carried out the most recent work, there are other botanical case reports on the successful micropropagation of A. auriculiformis [14],[16]. In black wattle (A. mearnsii), native plant from Australian, in vitro propagation was observed in nodal segments from 9-months-old seedlings in half-strength MS mineral salts supplemented with 0.05 or 0.1 mg L-1 BA and/or 0.05 mg L-1 IBA [17]. In A. nilotica subsp. indica, cotyledonary nodes differentiated multiple shoots in B5 medium [12] supplemented with the cytokinins BA, 2iP, KIN or ZEA, [18]. In A. nilotica subsp. hemispherica, an endangered and endemic species from Southern Pakistan, a micropropagation system was developed from seedlings in MS or B5 culture medium with various levels of BAP (0.5 to 4.0 mg L-1) along with 0.5 mg L-1 NAA, and MS medium produced the highest number of shoots[19]. In A. tortilis subsp. raddiana and A. nilotica, the micropropagation was observed only in nodal segments from 4-weeks-old seedlings of A. nilotica in MS culture medium with 2.5 mg L-1 BA, and rooting with 4.0 mg L-1 IBA [20]. Correnpondingly, in A. chundra, the bud sprout was obtained from shoot tip and nodal segment derived from seedlings on MS culture medium with 1.0 mg L-1 BA and 20 mg L-1 adenine sulfate [21].
On the other hand, studies and informations on Acacia micropropagation, using explants
obtained from juvenile plants under greenhouse conditions or from field plants,
are scarce. In A. mangium,
one of the most important forest species due to the quality of its wood, nodal
segments of three-month-old plants were used under greenhouse conditions,
observing that explants cultured in MS culture medium supplemented with 2.22 μM BAP and 2.69 μM NAA
induced the highest number of shoots and roots, respectively [22]. In another study conducted on A.
mangium and A.
mangium x A.
auriculiformis, an interspecific hybrid of
natural origin, micropropagation was started either from seeds or from explants
collected from outdoors on MS culture medium with 4.4 μM
BA [23]. In the last decade, some reviews articles have been published on in vitro work done on the Acacia species, such as the one carried
out by Beck and Dunlop [24], highlighting the excellent review carried out by Gantait et al.[25].
In other context, there are studies on
phytochemical aspects and biological activity of numerous Acacia species. In A. nilotica, methanolic leaf and bark extracts showed some
biological activity against Escherichia
coli and Candida glabrata,
respectively [26]. In A. arabica (A. nilotica),
phytochemical analysis showed the occurrence of several secondary compounds
such as tannins, terpenoids, anthraquinones, alkaloids, and others, and acetone
bark extract was found to be most potent against select MDR pathogenic bacteria
as Pseudomonas aeruginosa, E. coli, Streptococcus pyogenes, and others [27]. However, in a more complete study conducted on A. nilotica,
the raw extract of leaves using two methods (cold and hot) and three solvents
(butanol, ethanol and methanol), revealed the presence of alkaloids,
flavonoids, tannins, terpenoids, and others compounds, as long as the result of
antibacterial activity revealed more susceptibility toward Gram-positive
bacteria such as Streptococcus pyogenes
when it is compared to Gram-negative bacteria as Pseudomonas aeruginosa [28]. In another species of Acacia
such as A. saligna (A. cyanophylla),
leaves ethanol extracts showed antimicrobial activity against Klebsiella pneumoniae (Gram-negative)
and Micrococcus (Gram-positive),
and pathogenic fungi Aspergillus
fumigatus and C.
albicans, as long as the major phenol compounds
were gallic and p-Coumaric acids[29]. In this same species, water extract of flowers showed antifungal
and antibacterial activities against Penicillium
chrysogenum (3%) and Pectobacterium caratovorum
subsp. caratovorum
(MIC=100 μg/mL), respectively, and the most
abundant phenolic and flavonoid compounds were benzoic acid and naringenin,
respectively[30]. On the other hand, in Acacia
implexa and A.
falcata, native species of New South Wales, Australia, 70% aqueous ethanol
extracts showed antibacterial activity against sensitive and MDR strains of Staphylococcus aureus, and A. implexa
possessed high TPC (Total Phenolic Contents) and strong DPPH radical scavenging
abilities [31].
Considering this mentioned scenario, in this paper we report the establishment of culture conditions for plant propagation of microcuttings from adult trees of Vachellia macracantha (ex Acacia macracantha), an important woody species of the seasonally dry tropical forest of northern Peru.
2. MATERIALS AND METHODS
2.1. PLANTING STOCK
Three to five-old plants, on natural field conditions, were selected from located near Motupe–Olmos (Lambayeque, Peru) (Figure 1a, b and c). The shoot tip and nodal segment explants 50 mm were taken from the non-lignified part of young branches. All the leaves were removed, then the explants were dipped for 10 min and agitated in detergent solution and then in 0.2% Orthocide for 30 min. Subsequently the explants were surface-sterilized in 70% ethanol for 1 min and 5% commercial sodium hypochlorite solution (Clorox®) for 5 min before being rinsed three times in sterillized distilled water. The shoot and node segments were then cut in uninodal microcuttings of 10 to 15 mm in lenght and aseptically introduced into test tubes (18x150 mm) containing the culture medium. The same method was used on the micropropagation process.

Figure 1: Plant propagation and germplasm conservation of Vachellia macracantha (faique). a. Adult plants used as a living fence, b. Branches with flowers, c. Branches with fruits
2.2. CULTURE MEDIUM AND GROWNING CONDITIONS
For shoots elongation was used a medium consisting of the MS salts[32] and vitamins 1.0 mg L-1 thiamine.HCl and 100 mg L-1 myo-inositol. This basal medium was supplemented with NAA (0.0, 2.5, 5.0 and 7.5 mg L-1) in high concentration. In indeed, we have tested NAA (0.5 mg L-1) – BAP or KIN or 2iP (0.05, 0.5, 2.5, 5.0 and 10.0 mg L-1) interactions, respectively.
For shoots elongation the uninodal segments were cultured en MS medium supplemented with 0.02 mg L-1 NAA – 0.05, 0.5 or 2.5 mg L-1 BAP – 0.05 mg L-1 GA3, interaction. Rooting was tested in culture medium supplemented with NAA (0.05 to 2.0 mg L-1) or IBA (0.05 to 2.0 mg L-1).
Both culture medium, shoots elongation and rooting plants, containing 30 g L-1 sucrose was solidified with 7 g L-1 agar-agar and sterilized at 120 oC for 15 min. The pH was adjusted to 5.8±0.1 before autoclaving process. Explants were cultivated in pyrex tubes (18x150 mm) each containing 15 mL of culture medium. Cultures were maintained at 26±2 oC, under light (16 h day photoperiod; 35 μE.m-2. s-1) provided by cool fluorescent lamps (Grolux Sylvania).
After rooting, the micro-plants were carefully extracted from the growing tubes and were directly transplanted in pots on a substratum consisting of vermiculite mixed equally with river sand.
2.3. STATISTICAL ANALYSIS
Each experimemnt was repeated two times with 25 explants per replicate. All data were subjected to one-way analysis of variance test (ANOVA) and performed with version 3.4.0 R software. Tuckey’s test was used to comapre the mean values at the 5% level.
3. RESULTS
3.1. SHOOT ELONGATION
Growth responses of V. macracantha are summarized in tables 1, 2 and 3. Table 1 illustrates the effect of different concentrations of NAA on the shoot elongation and development of nodal segment explants. After 45 days of growth, nodal explants showed profuse callus formation on the basal region in contact with nutrient solution. Callus was observed in all treatments and the tissue turned brown after a few days.
Table 1: Effect of NAA on shoot development from three to five-year-old Vachellia macracantha plants, after 45 days of culture.
|
NAA (mg L-1) |
Survival rate (%) |
Shoot size (mm) |
Nodes formed (No) |
Leaves formed (No) |
Lateral branched (No) |
Defoliation (%) |
Rotting (%) |
|
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
|
2.5 |
100.0 |
35.0±16.9ª |
12.3 |
8.0 |
0.7 |
100.0 |
25.0 |
|
5.0 |
100.0 |
29.0±7.1c |
8.6 |
5.2 |
1.8 |
100.0 |
17.0 |
|
7.5 |
100.0 |
31.0±10.8b |
7.6 |
8.0 |
1.6 |
100.0 |
0.0 |
Data were
the mean ± SE.
Means within each column followed
by similar letters are not significantly different at 0.05 probability level.
Table 2: Effect of several cytokinins on shoot developmen from three to five-year-old Vachellia macracantha plants, after 45 days of culture.
|
Plant growth regulators (mg L-1) |
Response |
|||||
|
NAA |
BAP |
KIN |
2iP |
Callus induction (%) |
Shoot size (mm) |
Survival rate (%) |
|
0.5 |
0.05 |
|
|
100 (+) |
10.0 - 15.0 |
20.0 |
|
|
0.5 |
|
|
100 (+) |
10.0 - 15.0 |
20.0 |
|
|
2.5 |
|
|
100 (++) |
5.0 - 10.0 |
10.0 |
|
|
5.0 |
|
|
100 (++) |
5.0 - 10.0 |
0.0 |
|
|
10.0 |
|
|
100 (+++) |
< 5.0 |
0.0 |
|
0.5 |
|
0.05 |
|
100 (+) |
10.0 - 15.0 |
15.0 |
|
|
|
0.5 |
|
100 (+) |
10.0 - 15.0 |
15.0 |
|
|
|
2.5 |
|
100 (++) |
5.0 - 10.0 |
10.0 |
|
|
|
5.0 |
|
100 (++) |
5.0 - 10.0 |
10.0 |
|
|
|
10.0 |
|
100 (+++) |
< 5.0 |
0.0 |
|
0.5 |
|
|
0.05 |
100 (+) |
10.0 - 15.0 |
20.0 |
|
|
|
|
0.5 |
100 (+) |
10.0 - 15.0 |
15.0 |
|
|
|
|
2.5 |
100 (++) |
5.0 - 10.0 |
10.0 |
|
|
|
|
5.0 |
100 (++) |
5.0 - 10.0 |
10.0 |
|
|
|
|
10.0 |
100 (+++) |
< 5.0 |
0.0 |
Callus induction: +, callus < 5 mm Ø; ++, callus between 6-10 mm Ø; +++, callus > 10 mm Ø
Table 3: Effect of NAA-BAP-GA3 on shoot developmen from three to five-year-old Vachellia macracantha plants, after 45 days of culture.
|
Plant growth regulators (mg L-1) |
Response |
|||||||
|
NAA |
BAP |
GA3 |
Survival rate (%) |
Shoot size (mm) |
Nodes formed (No) |
Leaves formed (No) |
Defoliation (%) |
Rooting (%) |
|
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
0.0 |
|
0.02 |
0.05 |
0.05 |
100.0 |
31.5±13.5ª |
5.8 |
11.6 |
0.0 |
20.0 |
|
0.02 |
0.5 |
0.05 |
100.0 |
25.3±6.3b |
3.2 |
5.3 |
0.0 |
0.0 |
|
0.02 |
2.5 |
0.05 |
100.0 |
23.2±4.1c |
2.8 |
4.7 |
0.0 |
0.0 |
Data were the mean ± SE.
Means within each column followed by similar letters are not
significantly different at 0.05 probability level
The
treatment 2.5 mg L-1 NAA induced better shoot development than all of the other treatments and resulted in the formation of
a shoot which average 35 mm in length. The shoot showed twelve nodes and
produce three to four explants suitable to continue micropropagation. In this same
treatment defoliation was often observed in all plantlets but not all the
leaves fall. The root formation was only 25%. In the other treatments
conducted, 5.0 and 7.5 mg L-1 NAA, at the base of the nodal
segments, callus developed which had a greater volumen,
specially when using a concentration of 7.5 mg L-1
NAA. However, the proliferation of basal callus did not seem to affect the
development of the axillary bud. In these treatments, most of the explants
developed a shoot approximately 30 mm in length. The lateral branched was
increased from 0.7 to 1.8, and the defoliation was also 100% but with a dramatic leaves fall. Rooted plants was
observed only 5.0 mg L-1 NAA treatment with a rate estimated in 17%.
The
NAA-BAP, NAA-KIN and NAA-2iP treatments, also were tested on the shoot
initiation an elongation (Table 2), but the results not were
satisfactory. In all cases studied, after 45 days of the culture, shoots
reached a mean length of < 5 to 15 mm as compared to 35.0 mm (NAA 2.5 mg L-1)
or 31.5 mm (NAA-0.05 mg L-1 BAP-GA3 interaction). The precentage of explants with shoot greater than 10 mm in
length was only 20%, in some treatments, reaching the same survival rate.
In the
Table 3 the effect of the NAA-BAP-GA3 treatments were demostrated. In this case, the initiation and elongation of
the shoot was less than that obtained with NAA treatments, but the plants
constitution was more vigorous. A single shoot was formed the axillary bud and
it grew to produce three to six nodes suitable to continue micropropagation.
With respect to other parameters, basal callus formation and defoliation, not
were observed (Figure 1d). Rooted plants were observed only in the treatment
that incorporated 0.05 mg L-1 BAP with a rate estimated in 20%.
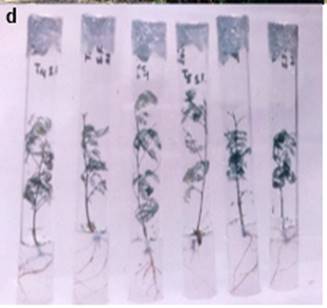
Figure 1: d. In vitro propagated plants
3.2. ROOT
INDUCTION AND ELONGATION
Root
induction was observed under several concentrations of either NAA or IBA
auxins. The Table 4 shows that for the NAA treatments, the root induction was
much less than that obtained with IBA. We have registered that in 0.5, 1.0 and
2.0 mg L-1 IBA treatments, after 30 days in culture, the root formed
were approximately 30 to 50 mm in length, more or less ramificated and very vigorous (Figure 1e). In this
culture medium formulation, although the defoliation rate was high, the
germplasm conservation was greater than 9 months. In this size (30 to 50 mm in lenght) rooted plantlets were transferred to a sterilized
substrate consisting of vermiculite and river sand, placed in a growth chamber
for one week, and later on transferred to the
greenhouse. Under these conditions survival rates were estimated over 80% since
V. macracantha
roots, average 3 to 4 per plant, showed normally rapid growth (Figure 1f).
Table 4: Effect of two auxins, NAA and IBA, on the root
formation in Vachellia macracantha micropropagated plants, after 45 days of culture.
|
Plant growth regulators (mg L-1) |
Response |
|||
|
NAA |
IBA |
Rooting (%) |
Root number |
Root size (mm) |
|
0.0 |
|
0.0 |
- |
- |
|
0.05 |
|
0.0 |
- |
- |
|
0.1 |
|
0.0 |
- |
- |
|
0.5 |
|
70.0 |
1-3 |
< 3.0 |
|
1.0 |
|
100.0 |
1-3 |
< 3.0 |
|
2.0 |
|
80.0 |
1-3 |
< 3.0 |
|
|
0.0 |
0.0 |
- |
- |
|
|
0.05 |
0.0 |
- |
- |
|
|
0.1 |
0.0 |
- |
- |
|
|
0.5 |
100.0 |
2-4 |
> 5.0 |
|
|
1.0 |
100.0 |
2-4 |
> 5.0 |
|
|
2.0 |
100.0 |
2-4 |
> 5.0 |
Root number: Main roots with numerous lateral
roots

Figure 1: e. In vitro plants stored for
more than a year, showing strong defoliation, and f. Plants acclimatization
4. DISCUSSION
In the
micropropagation of Acacia species,
studies have been started almost entirely with seedlings of different ages,
with the exception of the study carried out in A. albida by Gassama and Duhoux [8], similar findings showed in A. senegal by Badji et al.[9] and in the present study carried out in V. macracantha.
Likewise, only in the study carried out in A.
albida by Duhoux and Davies [10], in the micropropagation process, 0.5 mg L-1
NAA - 3.0 mg L-1 BAP was used. However, in pioneering studies
conducted on Fabaceae species, high levels of NAA (3.0 to 10.0 mg L-1)
were most efficient for rooting and propagation of plantlets in several species
of Prosopis (P. chilensis, P. alba and
P. tamarugo)[5],[33]. These authors considered that NAA is an
only hormone required to trigger root initiation and that the addition of GA3,
although it is inhibitory to some extent, favoured
axillary buds growing. This result was considered of extreme importance,
since explant normally shown leaf abscission after some days of culture
and axillary buds do not initiate growth immediately after root formation[33]. In our system, all concentrations of NAA
tested were observed both, axillary bud development and leaf abscission. In
others species of Acacia as A. senegal [9], A. koa [34], A. auriculiformis [13], A. nilotica [20], and others, NAA not was tested in the plant
propagation process.
In the
micropropagation (elongation and shoot proliferation) of numerous Acacia species, cytokinins
as N6-benzyladenine (BA), 6-furfurylaminopurine (KIN), N6-(2-isopentenyl)
adenine (2iP), 4-hydroxy-3-methyl-terms-2-butenyl aminopurine (ZEA) and 1-phenyl-3-
(1,2,3-thiadiazol-5-yl) urea (TDZ), have been the main growth regulators
used and/or tested in several experiments. In our study, only some interactions
of NAA-BAP, NAA-KIN and NAA-2iP, in the lowest concentrations tested, showed little
shoot elongation (≤ 15 mm), although with a survival rate not more than
20%. However, in most studies the BA supplement has shown better results than
the other cytokinins, as reported in A. nilotica
subsp. indica [18], A. nilotica [20], A. mearnsii [17], A chundra [21] and A.
auriculiformis [13]. In another investigations, it has been
shown that the culture medium containing BA was much superior to the culture
medium containing KIN, for shoot multiplication of A. mangium [35],[36], and Badji et al.[9]reported that ZEA produced better induction
to multiple shoot formation in A. senegal. However, in A. nilotica the induction of multiple
shoots was observed when 0.6 mg L-1 NAA and it was supplemented to
the culture medium, one month after that the explant was cultivated with 1.0 mg
L-1 KIN [37]. On the other hand, in V. macracantha the best results in the
shoot elongation were observed in various combinations of NAA-BAP-GA3.
In this way, there are some studies where the use of a single growth regulator
was not as efficient as when combinations of BA-NAA or TDA-NAA were used, as
observed in the shoot’s proliferation of A.
seyal [38]. This first observation was ratified with
the experiments conducted by Rout et al. [21] regarding in A. chundra, who have used the BA-IAA
interaction, by Abbas et al.[19] associated A. nilotica subsp. hemispherica, using the BAP-NAA
interaction, Shahinozzaman et al.[36] in A. mangium, using the BAA-NAA interaction and in Yadav et
al. [39] in A. auriculiformis, using the KIN-IAA interaction. In
general, in V. macracantha
BAP, a sintetic cytokinin, in interaction with auxin
(NAA), produced a better stimulation of the shoot growth and a vigorous
plantlets formation, that using cytokinin alone, as postulated by Gantait et al.[25].
In our
findings regarding V. macracantha,
we have showed that various concentrations of NAA and IBA are directly related with the root formation. However, in
IBA treatments the percentage of rooted plants reached 100%, which is always
more difficult to induce when non-juvenile structures are used as explants [25]. IAA, IBA or NAA auxins have been used in
numerous studies on in vitro rooting
of Acacia, in various concentrations
and generally individually [13],[40][41]. Likewise, as observed in our
study in V. macracantha,
IBA has been more successful than IAA or NAA [35],[42].
5. CONCLUSIONS
Vachellia macracantha (“faique”), Prosopis limensis (“algarrobo”)
and Colicodendron escabridum
(“zapote”), are the most representative forest species of the Seasonally Dry
Tropical Forest of northern Peru, as well as those of greater economic
importance. That is why this species have been subjected to strong anthropic
pressure that the best genotypes have now been exterminated. Given this, the
development of biotechnological methods, such as in vitro tissue culture, has allowed its propagation from young
specimens, achieving elongation of the shoot in culture medium supplemented
with NAA or NAA-BAP-GA3 and plantlets rooting in culture medium
supplemented with IBA. These results will enable the spread of V. macracantha
on a large scale, contributing to the reforestation of the species and the
mitigation of climate change in northern Peru.
SOURCES OF FUNDING
This worked has financied by the General Biotechnology Laboratory – VRINV (Universidad Nacional Pedro Ruiz Gallo).
CONFLICT OF INTEREST
None.
ACKNOWLEDGMENT
Our thanks to Pilar Bazán-Sernaqué, Boris Esquerre-Ibañez
and Héctor Valdiviezo-Castillo for preparing the
figures.
REFERENCES
[1]
Maslin,
B. R., Miller, J. T. and Seigler, D. S. (2003): Overview of the generic status of Acacia (Leguminosae: Mimosoideae). Australian Systematic Botany, 16: 1-18.
[2]
Galiana, A., Balle, P., Kanga, N’. G. and Domenach, A. M. (2002): Nitrogen fixation estimated by the
15N natural abundance method in Acacia mangium Willd. inoculated with Bradyrhizobium
sp. and grown in sylvicultural conditions. Soil Biology and Biochemistry, 34:
251-262.
[3]
Soukup,
J. (1970): Vocabulario de los nombres vulgares
de la flora peruana. Edit. Colegio Salesiano, Lima,
Perú. 384 p.
[5]
Jordan,
M., Pedraza, J. and Goreux, A. (1985): In vitro propagation studies of three Prosopis species (P.
alba, P. chilensis and P. tamarugo) through shoot tip
culture. Gartenbauwiossenschaft, 50: 265-267.
[6]
Felker,
P., Canell, G. H., Clark, P. R., Osborn, J. F. and Nash, P. (1983): Biomass
production of Prosopis species (mesquite), Leucaena and others leguminous trees
grown under heat/drougt stress. Forest Science, 29:
592-606.
[8]
Gassama,
Y. K. and Duhoux, E. (1986): Micropropagation of l’ Acacia albida Del. (Leguminosae) adulte. Bulletin de l’IFAN Cheik Anta Diop, T.46, serie A,
34: 314-320.
[9]
Badji, S., Mairone, Y., Ndiaye, I., Merlin, G., Danthu, P., Neville, P. and Colonna, J. P. (1993): In vitro
propagation of the gum arabic tree (Acacia senegal (L.) Willd.). 1. Developing
a rapid method for producing plants. Plant Cell Reports, 12: 629-633.
[10] Duhoux, E. and Davies, D. (1985): Caulogénèse è partir des bourgeons cotylédonaires
d’ Acacia albida et influence du saccharose sur la rhizogénèse.
Journal of Plant Physiology, 121: 175-180.
[11] Lloyd, G. and McCown, B. (1981): Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot
tip culture. Combined Proceedings – International Plant Propagator’s Society,
30: 421-427.
[12] Gamborg, O. L., Miller, R. A. and Ojima, K. (1968):
Nutrient requeriments of suspensión
cultures of soybean root cells. Experiment Cell Research, 50: 151-158.
[13] Ismail, H., Muniando, S. K., Yusoff, A. M., Hassan, N. H. and Ab Shukor, N. A. (2016): In vitro micropropagation of Acacia auriculiformis from selected juvenile sources. Dendrobiology, 75: 157-165.
[14] Mittal, A., Agarwal, R. and Gupta,
S. C. (1989): In vitro development of plantlets from axillary buds of Acacia auriculiformis – a leguminous tree. Plant Cell, Tissue and Organ Culture, 19: 65-70.
[15]
Girijashankar, V. (2011): Micropropagation of multipurpose
medicinal tree Acacia auriculiformis. Journal of
Medicinal Plant Research, 5: 462-466.
[16] Banerjee, P. (2013): Rapid in
vitro propagation of Acacia auriculiformis on solid and liquid media:role of organic additive,
antioxidant and plant growth regulators. Cibtech
Journal of Bio-Protocols, 2: 39-49.
[17] Correia, D. and Graça, M. E. C. (1995): In vitro propagation of black wattle (Acacia meamsii De Wild). IPEF, 48/49: 117-125.
[18] Dewan, A., Nanda, K. and Gupta, S.
C. (1992): In vitro micropropagation of Acacia nilotica subsp. indica Brenan via
cotyledonary nodes. Plant Cell Reports, 12: 18-21.
[19] Abbas, H., Qaiser, M. and Naqvi, B. (2010): Rapid in vitro multiplication of Acacia nilotica subsp. hemispherica, a
critically endangered endemic taxo. Pakistan Journal
of Botany, 42: 4087-4093.
[20] Aziz, A. A. S., Omari, M. A. and Kafawin, O. M. (2002): Micropropagation of Acacia tortilis
subsp. raddiana and A. nilotica
under in vitro conditions. Journal of Tropical Forest Science, 14: 329-336.
[21] Rout, G. R., Senapati, S. K. and Aparajeta, S. (2008): Micropropagation of Acacia chundra
(Roxb.) DC. Horticultural Science (Prague), 35:
22-26.
[22] Torres, L. A., Suárez, I. E. and Gatti, K. (2013): Propagación in vitro de Acacia mangium Willd. Biotecnología en el Sector Agropecuario y Agroindustrial,
11: 81-87.
[23] Monteuuis, O., Galiana, A. and Goh, D. (2013): In
vitro propagation of Acacia mangium and A. mangium x A. auriculiformis. In:
Protocols for Micropropagation of Selected Economically-Important
Horticultural Plants. Methods in Molecular Biology. M. Lambardi
et. al. (eds.). Vol. 994, pp. 199-211. Springer Science-Business Media New
York.
[25] Gantait, S., Kundu, S. and Das, P. K. 2018. Acacia: An exclusive survey on in
vitro propagation. Journal of the Saudi Society of Agricultural Sciences, 17:
163-177.
[26] Dev, S. N. C., De, K. and Singh,
S. (2014): Antimicrobial activity and phytochemical analysis of Acacia nilotica (L.) Del. Indian Journal Applied & Pure Biology, 29: 331-332.
[27] Lawrence, R., Jeyakumar, E. and Gupta, A. (2015): Antibacterial activity of Acacia arabica
(bark) extract against selected multi drug resistant pathogenic bacteria.
International Journal of Current Microbiology and Applied Sciences, Special
Issue-1: 213-222.
[28] Singh, R. and Thakur, R. (2016):
Phytochemical analysis and antibacterial activity of Acacia nilotica (L.) leaves against pathogenic bacteria. International Journal of
Green Pharmacy, 10: 104-110.
[29] Gumgumjee, N. M. and Hajar, A. S. (2015): Antimicrobial efficacy of Acacia saligna (Labill.) H.L. Wendl. and Cordia sinensis Lam. leaves extracts against
some pathogenic microorganisms. International Journal of Microbiology and
Immunology Research, 3: 51-57.
[30] Al-Huqail, A. A., Behiry,
S. I., Salem, M. Z. M., Ali, H. M., Siddiqui, M. H. and Salem, A. Z. M. (2019):
Antifungal, antibacterial, and antioxidant activities of Acacia saligna (Labill.) H.L. Wendl. flower extract: HPLC analysis of phenolic and
flavonoid compounds. Molecules, 24, 700.
[31] Akter, K., Barnes, E. C., Brophy, J. J., Harrington, D., Elders, Y. C., Vemulpad, S. R. and Jamie, J. F. (2016): Phytochemical
profile and antibacterial and antioxidant activities of medicinal plants used
by aboriginal people of New South Wales, Australia. Evidence-based
Complementary and Alternative Medicine, Volume 2016, Article ID 4683059, 14
pages.
[32] Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with
tobacco tissue cultures. Physiologie Plantarum 15:
473-497.
[33] Jordan, M. and Balboa, O. (1985):
In vitro regeneration of Prosopis tamarugo Phil. and Prosopis chilensis
(Mol.) Stuntz from nodal sections. Gartenbauwissenschaft, 50: 138-142.
[34] Skolmen, R. G. (1986). Acacia (Acacia koa Grey). In:
Biotechnology in Agriculture and Forestry. Y. P. S. Bajaj (ed.). Vol. 1: Trees
I, Springer-Verlag, Berlin. 375-384 pp.
[35] Shahinozzaman, M., Azad, M. A. K. and Amin, M. N. (2012): In vitro clonal
propagation of a fast-growing legume tree-Acacia mangium
Willd. employing cotyledonary node explants. Notulae Scientiae Biologicae, 4:
79-85.
[36] Shahinozzaman, M., Farug, M. O., Ferdous, M. M., Azad, M.
A. K. and Amin, M. N. (2013): Direct organogenesis and plant regeneration from
cotyledons of a multipurpose tree, Acacia mangium Willd. Current Trends in Biotechnology and Pharmacy, 7:
511-517.
[37] Dhabhai, K., Sharma, M. M. and Batra, A. (2010): In vitro clonal propagation
of Acacia nilotica (L.) – a nitrogen fixing tree.
Researcher, 2: 7-11.
[38] Al-Wasel, A. S. (2000): Micropropagation
of Acacia seyal Del. in vitro. Journal of Arid
Environment, 46: 425-431.
[39] Yadav, R., Yadav, N. and Kumar, S.
(2015): An improved micropropagation and assessment of genetic fidelity in
multipurpose medicinal tree Acacia auriculiformis. Proceedings of the National
Academic of Sciences, India Section B: Biological Sciences, 86: 921-929
[40] Khalafalla, M. M. and Daffalla, H. M. (2008): In vitro
micropropagation and micrografting of gum Arabic tree [Acacia senegal (L.) Willd].
International Journal of Sutainable Crop Production,
3: 19-27.
[41] Rathore, J. S., Rai, M. K., Phulwaria, M. and Shekhawat, N. S. (2014): A liquid
culture system for improvemed micropropagation of
mature Acacia nilotica (L.) Del. spp. indica and ex
vitro rooting. Proceedings of the National Academic of Sciences, India Section
B: Biological Sciences, 84: 193-200.
[42] Javed, S. B., Anis, M., Khan, P. R. and Aref, I.
M. (2013): In vitro regeneration and multiplication for mass propagation of
Acacia ehrenbergiana Hayne: a potential reclaiment of denude arid lands. Agroforestry Systems, 87:
621-629.
|
|
 This work is licensed under a: Creative Commons Attribution 4.0 International License
This work is licensed under a: Creative Commons Attribution 4.0 International License
© Granthaalayah 2014-2020. All Rights Reserved.